
Triceratops

| Triceratops | |
|---|---|

| |
| ’n Triceratops-skelet (Natuurhistoriese Museum van Los Angeles). | |
| Wetenskaplike klassifikasie | |
| Koninkryk: | |
| Filum: | |
| Klas: | |
| Superorde: | |
| Orde: | |
| Infraorde: | |
| Familie: | |
| Genus: | Triceratops (Marsh, 1889)
|
| Spesies | |
| |

| Triceratops | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Kladistiese klassifikasie | ||||||||||||||
|
Triceratops, of die triseratopse of trikeratopse (Grieks vir "driehoringgesig"), is ’n genus van plantvretende dinosourusse wat in die Laat Maastricht-tydsnede van die Laat Kryt-periode, sowat 68 miljoen jaar gelede, vir die eerste keer voorgekom het in wat nou Noord-Amerika is. Dit is een van die laaste bekende nie-vlieënde dinosourus-genera en het in die Kryt-Paleogeen-uitwissing 66 miljoen jaar gelede uitgesterf.[1]
Die triseratops het ’n groot, benerige kraag en drie horings op sy groot, viervoetige lyf gehad en het nogal soos die moderne renoster gelyk. Dit is een van die herkenbaarste dinosourusse en die bekendste seratopside. Dit het saam met die vreesaanjaende Tyrannosaurus bestaan en is waarskynlik deur laasgenoemde gejag,[2] hoewel dit minder seker is of die twee wel baklei het soos in tradisionele museums en gewilde prente uitgebeeld word.
Twee spesies, T. horridus en T. prorsus, word erken, hoewel baie ander spesies ook al genoem is. Volgens navorsing wat in 2010 gepubliseer is, is Torosaurus, ’n seratopside wat lank as ’n aparte genus beskou is, ’n Triceratops,[3][4] maar nie alle wetenskaplikes aanvaar dit nie.[5][6]
Beskrywing[wysig | wysig bron]
Individuele triseratopse het na raming lengtes van 7,9 tot 9 m gehaal en hoogtes van 2,9 tot 3 m,[7][8] en kon 6,1 tot 12 ton geweeg het.[9] Hul grootste kenmerk was hul groot skedel; dit was van die grootste onder alle landdiere. Die grootste bekende skedel (monster BYU 12183) was na raming 2,5 m lank[3] en kon tot sowat ’n derde van die dier se hele lengte uitgemaak het.[10] Dit het ’n enkele horing op die neus gehad, net bo die neusgate, asook ’n horing van sowat 1 m bo elke oog.[11] Aan die agterkant van die skedel was ’n relatief kort, benerige kraag, met skerp punte by sekere spesies. Die meeste ander seratopsides het groot openinge in die kraag gehad, terwyl die triseratops s’n solied was.[12]
Die dier se vel was ongewoon in vergelyking met ander dinosourusse s’n. Velafdrukke van ’n tot nog toe onbeskrewe fossiel wys sommige spesies was dalk bedek met haaragtige stekels, soos dié van die meer primitiewe seratopsiër Psittacosaurus.[13]
Ledemate[wysig | wysig bron]
Triseratops-spesies was stewig gebou en het sterk ledemate gehad, asook kort hande met drie hoewe elk, en kort voete met vier hoewe elk.[14] Hoewel hulle beslis vierpotiges was, word al lank gedebatteer oor die diere se houding. Oorspronklik is geglo die voorpote was uitgesprei om die gewig van die kop beter te kon dra.[15] Gefossileerde voetspote van dinosourusse met horings en onlangse rekonstruksies van skelette (beide fisiek en digitaal) dui daarop triseratops en ander seratopsides het ’n regop houding gehad as hulle loop, met die elmboë gebuig – ’n houding tussen heeltemal regop en met die pote uitgesprei soos by moderne renosters.[14][16][17][18]
Die hande en voorarms van die triseratops het ’n taamlik primitiewe struktuur gehad in vergelyking met ander vierpotige dinosourusse soos die Thyreophora-spesies en baie souropodes. In dié twee groepe was die voorpote van vierpotige spesies gewoonlik gedraai sodat die hande vorentoe en die palms agtertoe gewys het. Triseratopse het, nes ander seratopsiërs en die verwante vierpotige ornitopodes, geloop met hul vingers wat weg van die lyf na buite wys – die primitiewe manier wat ook aangetref is by tweepotige dinosourusse soos teropodes.
Klassifikasie[wysig | wysig bron]
Triceratops is die bekendste genus van Ceratopsidae, ’n familie groot Noord-Amerikaanse Ceratopsia-dinosourusse met horings. Daar is egter lank gedebatteer oor die presiese plasing van Triceratops in die familie. Verwarring word veral veroorsaak deur die kombinasaie van kort, soliede krae (soorgelyk aan dié van Centrosaurinae) en die lang horings bo die oë (meer soos dié van Ceratopsidae, ook bekend as Chasmosaurinae).[19] In die eerste oorsig van gehoringde dinosourusse het die Amerikaanse paleontoloog R.S. Lull twee teorieë oor moontlike linies gehad: een van Monoclonius en Centrosaurus, wat tot Triceratops gelei het, en die ander van Ceratops en Torosaurus, wat Triceratops ’n sentrosourine maak soos wat die groep vandag verstaan word.[20] Latere hersienings het dié mening ondersteun en die eerste groep, met kort krae, beskryf as Centrosaurinae (insluitende Triceratops), en die tweede groep, met lang krae, as Chasmosaurinae.[21][22]
In 1949 het die Amerikaans-Kanadese paleontoloog C.M. Sternberg eerste dié teorie bevraagteken; hy het gemeen Triceratops was nader verwant aan Arrhinoceratops en Chasmosaurus, gebaseer op hul skedels en horings. Dit het Triceratops ’n genus van seratopsine (hy het dit chasmosourine genoem) gemaak.[23] Die meeste ander wetenskaplikes, soos John Ostrom[24] en later David Norman, het Triceratops in Centrosaurinae geplaas.[25]
Daaropvolgende ontdekkings en ontledings het Sternberg se mening oor Triceratops ondersteun. Lehman het albei subfamilies in 1990 gedefinieer en Triceratops as ’n seratopsine (chasmosourine) gereken op grond van verskeie morfologiese eienskappe. Buiten die kort kraag, pas dit baie goed in die seratopsine-subfamilie.[26] Verdere navorsing deur die Amerikaner Peter Dodson, onder meer ’n kladistiese ontleding in 1990,[27] het Triceratops se plasing in die seratopsine-subfamilie verder ondersteun.
Hier onder is ’n kladogram volgens Sampson et al. (2010).[28]

| Ceratopsidae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Gebruik in filogenetika[wysig | wysig bron]
In filogeniese taksonomie word die genus gebruik as ’n verwysingspunt in die definisie van Dinosauria; dinosourusse word gereken as alle afstammelinge van die mees onlangse gemeenskaplike voorsaat van Triceratops en Neornithes (dus moderne voëls).[29] Verder word die dinosourusse "met voëlheupe", Ornithischia, gedefinieer as alle dinosourusse met ’n meer onlangse gemeenskaplike voorsaat met Triceratops as met moderne voëls.[30]
Paleobiologie[wysig | wysig bron]

Hoewel triseratopse gewoonlik uitgebeeld word as diere wat in troppe voorgekom het, is daar tans min getuienis dat dit die geval was. Terwyl verskeie ander genera van gehoringde dinosourusse op terreine ontdek is met van twee tot honderde of duisende individue, is daar tot nou net een aangetekende geval van ’n terrein wat deur triseratops-bene oorheers word: die oorblyfsels van drie jong diere is in Montana gevind. Dit is dalk betekenisvol dat al drie diere jonk was.[31]
’n Ander, meer onlangse vonds kan daarop dui dat triseratopse wel in klein familiegroepe voorgekom het. In 2012 is drie triseratops-individue in ’n relatief volledige toestand in Wyoming ontdek. Hul grootte het gewissel van ’n volgroeide dier tot ’n baie jong een. Die oorblyfsels word tans deur paleontoloë ondersoek. Daar word geglo die diere het as ’n gesinseenheid gereis, maar dit is nog onseker of hulle ’n paar en hul kleintjie was, of twee wyfies wat na ’n jong dier omgesien het. Die oorblyfsels, veral van die grootste individu, toon ook tekens dat dit óf aangeval óf agterna geaas is deur ’n Tyrannosaurus; die bene van die voorpote is gebreek en het tandmerke van ’n Tyrannosaurus aan.[32]
Individuele fossiele van triseratopse word al baie jare lank gevind.[31] Dié oorblyfsels is baie algemeen; die paleontoloog Bruce Erickson het byvoorbeeld berig dat hy 200 fossiele in Montana gevind het.[33] Barnum Brown beweer hy het meer as 500 skedel daar gevind.[15] Omdat fossiele tande en dele van horings, krae en skedels uit die Laat Maastricht-tydsnede van die Laat Kryt-periode, 66 miljoen jaar gelede, in westelike Noord-Amerika so volop is, word dit beskou as een van die algemeenste plantvreters van sy tyd, indien nie die algemeenste een nie. Die Amerikaanse paleontoloog Robert Bakker het in 1986 geraam dat vyf sesdes van die groot dinosourusse aan die einde van die Kryt triseratopse was.[34]
Triceratops was een van die laaste seratopsiërs wat sy verskyning gemaak het voor die Kryt-Paleogeen-uitwissing. Die verwante Torosaurus en die verder verwante klein Leptoceratops was ook teenwoordig, hoewel min fossiele van hulle ontdek is.[15]
Tande en dieet[wysig | wysig bron]

Triseratopse was plantvreters, en omdat hul koppe laag was, het hulle meestal laaggeleë plante geëet. Hulle kon ook hoër plante met hul horings, snoet en lyf platgeslaan het.[35][36] Die bek lyk of dit beter kon gryp en pluk as byt.[24]
Die diere se tande was gerangskik in 36 tot 40 kolomme aan elke kant van elke kaak, met drie tot 3 tot 5 tande per kolom, na gelang van die individu se grootte.[35] Hulle het dus van 432 tot 800 tande gehad, waarvan net ’n deel op ’n slag gebruik is (tande is voortdurend deur die dier se lewe gewissel).[35] Hulle het plantmateriaal in ’n vertikale rigting geskeur.[35] Die grootte en getal tande dui daarop dat hulle groot hoeveelhede veselryke plante geëet het,[35] dalk palms en broodbome,[37][38] of dalk varings.[39]
Doel van die horings en kraag[wysig | wysig bron]
Daar is al baie gegis oor die doel van die triseratops se horings en kraag. Die twee hoofteorieë is dat dit gebruik is vir gevegte of om te pronk voor ’n lid van die teenoorgestelde geslag.[35]
Vroeër is gemeen die kraag het dalk gedien as ankerpunte vir die kaakspiere om die dier beter te laat kou.[40] In latere studies is egter geen getuienis gevind van groot spieraanhegsels aan die kraagbene nie.[41]
Die groot kraag kon ook die lyf se oppervlak vergroot het om liggaamshitte te help reguleer.[42] Dié doel alleen sou egter nie die bisarre en uitspattige vorms verduidelik wat in verskillende lede van Ceratopsidae te bespeur is nie.[35]
Die teorie dat hulle gebruik is in seksuele vertoon, is in 1961 die eerste keer deur Davitashvili voorgestel en word sedertdien al hoe meer aanvaar.[26][41][43] Gehoringde dinosourusse se pantsers verskil aansienlik en dit ondersteun die vertoonteorie omdat dit elke spesie besonders maak. Verder gebruik moderne diere met soortgelyke versierings dit ook vir seksuele vertoon.[44]
Verwysings[wysig | wysig bron]
- ↑ Scannella, J.B., & Fowler, D.W. (2014). "A stratigraphic survey of Triceratops localities in the Hell Creek Formation, northeastern Montana (2006–2010)". Geological Society of America Special Papers. 503: 313–332. doi:10.1130/2014.2503(12).
{{cite journal}}: AS1-onderhoud: gebruik authors-parameter (link) - ↑ Erickson, GM; Olson, KH (1996). "Bite marks attributable to Tyrannosaurus rex: preliminary description and implications". Journal of Vertebrate Paleontology. 16 (1): 175–178. doi:10.1080/02724634.1996.10011297.
- ↑ 3,0 3,1 Scannella, J.; Horner, J.R. (2010). "Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae): synonymy through ontogeny". Journal of Vertebrate Paleontology. 30 (4): 1157–1168. doi:10.1080/02724634.2010.483632.
- ↑ Switek, Brian. "New Study Says Torosaurus=Triceratops". Dinosaur Tracking (in Engels). Smithsonian.com. Geargiveer vanaf die oorspronklike op 13 November 2013. Besoek op 2 Maart 2011.
- ↑ Farke, Andrew A. (2011). Claessens, Leon (red.). "Anatomy and taxonomic status of the chasmosaurine ceratopsid Nedoceratops hatcheri from the Upper Cretaceous Lance Formation of Wyoming, U.S.A". PLoS ONE. 6 (1): e16196. doi:10.1371/journal.pone.0016196. PMC 3024410. PMID 21283763.
- ↑ [1], Longrich NR, Field DJ (2012) Torosaurus Is Not Triceratops: Ontogeny in Chasmosaurine Ceratopsids as a Case Study in Dinosaur Taxonomy. PLoS ONE 7(2): e32623. doi:10.1371/journal.pone.0032623
- ↑ "T Dinosaurs Page 2" (in Engels). DinoDictionary.com. Geargiveer vanaf die oorspronklike op 11 Mei 2020. Besoek op 3 Augustus 2010.
- ↑ "Triceratops in The Natural History Museum's Dino Directory" (in Engels). Internt.nhm.ac.uk. Geargiveer vanaf die oorspronklike op 17 Februarie 2009. Besoek op 3 Augustus 2010.
- ↑ Alexander, R.M. (1985). "Mechanics of posture and gait of some large dinosaurs". Zoological Journal of the Linnean Society. 83: 1–25. doi:10.1111/j.1096-3642.1985.tb00871.x.
- ↑ Lambert, D. (1993). The Ultimate Dinosaur Book. Dorling Kindersley, New York. pp. 152–167. ISBN 1-56458-304-X.
- ↑ "Denver museum unveils 7-foot-long, 1,000-pound Triceratops skull". The Daily Courier. 18 November 2003. Besoek op 26 Desember 2013.
- ↑ "Making A Triceratops .Science Supplies Missing Part! Of Skeleton". Boston Evening Transcript. 24 Oktober 1901. Besoek op 26 Desember 2013.
- ↑ Perkins, S.; Csotonyi, Julius T. (2010). "Dressing Up Dinos". Science News. 177 (3): 22–25. doi:10.1002/scin.5591770321. Geargiveer vanaf die oorspronklike op 29 September 2012. Besoek op 22 April 2014.
- ↑ 14,0 14,1 Fujiwara, S.-I. (2009). "A Reevaluation of the manus structure in Triceratops (Ceratopsia: Ceratopsidae)". Journal of Vertebrate Paleontology. 29 (4): 1136–1147. doi:10.1671/039.029.0406.
- ↑ 15,0 15,1 15,2 Dodson, P. (1996). The Horned Dinosaurs. Princeton University Press, Princeton, New Jersey. ISBN 0-691-02882-6.
- ↑ Christiansen, P.; Paul, G.S. (2001). "Limb bone scaling, limb proportions, and bone strength in neoceratopsian dinosaurs" (PDF). Gaia. 16: 13–29.
- ↑ Thompson, S.; Holmes, R. (2007). "Forelimb stance and step cycle in Chasmosaurus irvinensis (Dinosauria: Neoceratopsia)". Palaeontologia Electronica. 10 (1): 17 p.
- ↑ Rega, E.; Holmes, R.; Tirabasso, A. (2010). "Habitual locomotor behavior inferred from manual pathology in two Late Cretaceous chasmosaurine ceratopsid dinosaurs, Chasmosaurus irvinensis (CMN 41357) and Chasmosaurus belli (ROM 843)". New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium. Bloomington and Indianapolis: Indiana University Press. pp. 340–354. ISBN 978-0-253-35358-0.
{{cite book}}: Onbekende parameter|editors=geïgnoreer (hulp) - ↑ "What is special about the Triceratops?" (in Spaans). Dinosaurios.org. Geargiveer vanaf die oorspronklike op 31 Mei 2020. Besoek op 26 Desember 2013.
- ↑ Hatcher, J. B., Marsh, O. C., and Lull, R. S. (1907) The Ceratopsia. Government Printing Office, Washington, D.C. ISBN 0-405-12713-8.
- ↑ Lambe, L.M. (1915). "On Eoceratops canadensis, gen. nov., with remarks on other genera of Cretaceous horned dinosaurs". Canada Department of Mines Geological Survey Museum Bulletin. 12: 1–49. ISBN 0-665-82611-7. Geargiveer vanaf die oorspronklike op 29 Junie 2011. Besoek op 22 April 2014.
- ↑ Lull, R. S. (1933). "A revision of the Ceratopsia or horned dinosaurs". Memoirs of the Peabody Museum of Natural History. 3 (3): 1–175. Besoek op 20 November 2010.
- ↑ Sternberg, C. M. (1949). "The Edmonton fauna and description of a new Triceratops from the Upper Edmonton member; phylogeny of the Ceratopsidae". National Museum of Canada Bulletin. 113: 33–46.
- ↑ 24,0 24,1 Ostrom, J. H. (1966). "Functional morphology and evolution of the ceratopsian dinosaurs". Evolution. 20 (3): 290–308. doi:10.2307/2406631. JSTOR 2406631.
- ↑ Norman, David (1985). The Illustrated Encyclopaedia of Dinosaurs. London: Salamander Books. ISBN 0-517-46890-5.
- ↑ 26,0 26,1 Lehman, T. M. (1990). The ceratopsian subfamily Chasmosaurinae: sexual dimorphism and systematics. in: Carpenter, K., and Currie, P. J. (reds.). Dinosaur Systematics: Perspectives and Approaches. Cambridge University Press, Cambridge, pp. 211–229. ISBN 0-521-36672-0.
- ↑ Dodson, P., and Currie, P. J. (1990). Neoceratopsia. 593–618. in Weishampel, D. B., Dodson, P., & Osmólska, H. (reds.). The Dinosauria. University of California Press, Berkeley, pp. 593–618. ISBN 0-520-06727-4.
- ↑ Scott D. Sampson, Mark A. Loewen, Andrew A. Farke, Eric M. Roberts, Catherine A. Forster, Joshua A. Smith, and Alan A. Titus (2010). "New Horned Dinosaurs from Utah Provide Evidence for Intracontinental Dinosaur Endimism". PLoS ONE. 5. 5 (9): e12292. doi:10.1371/journal.pone.0012292. PMC 2929175. PMID 20877459.
{{cite journal}}: AS1-onderhoud: gebruik authors-parameter (link) - ↑ Gauthier, J. A. (1986). "Saurischian monophyly and the origin of birds. The Origin of Birds and the Evolution of Flight, K. Padian (red.)". Memoirs of the California Academy of Sciences. 8: 1–55.
- ↑ Sereno, P. C. (1998). "A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 210 (1): 41–83.
- ↑ 31,0 31,1 Mathews, Joshua C.; Brusatte, Stephen L.; Williams, Scott A.; Henderson, Michael D. (2009). "The first Triceratops bonebed and its implications for gregarious behavior". Journal of Vertebrate Paleontology. 29 (1): 286–290. doi:10.1080/02724634.2009.10010382.
- ↑ Triceratops trio unearthed in Wyoming. CNN.com. Besoek op 2013-08-25.
- ↑ Erickson, B.R. (1966). "Mounted skeleton of Triceratops prorsus in the Science Museum". Scientific Publications of the Science Museum. 1: 1–16.
- ↑ Bakker, R.T. (1986). The Dinosaur Heresies: New Theories Unlocking The Mystery of the Dinosaurs and Their Extinction. William Morrow, New York. ISBN 0-14-010055-5.
- ↑ 35,0 35,1 35,2 35,3 35,4 35,5 35,6 Dodson, P.; Forster, C.A.; en Sampson, S.D. (2004) Ceratopsidae. In: Weishampel, D. B.; Dodson, P.; and Osmólska, H. (reds.), The Dinosauria (2de uitg.) University of California Press, Berkeley, pp. 494–513. ISBN 0-520-24209-2.
- ↑ Tait, J.; Brown, B. (1928). "How the Ceratopsia carried and used their head". Transactions of the Royal Society of Canada. 22: 13–23.
- ↑ Ostrom, J. H. (1964). "A functional analysis of jaw mechanics in the dinosaur Triceratops" (PDF). Postilla. 88: 1–35. Geargiveer vanaf die oorspronklike (PDF) op 26 Junie 2010. Besoek op 20 November 2010.
- ↑ Weishampel, D. B. (1984). "Evolution of jaw mechanisms in ornithopod dinosaurs". Advances in Anatomy, Embryology, and Cell Biology. 87: 1–110. PMID 6464809.
- ↑ Coe, M. J.; Dilcher, D. L.; Farlow, J. O.; Jarzen, D. M.; and Russell, D. A. (1987). Dinosaurs and land plants. In: Friis, E. M.; Chaloner, W. G.; and Crane, P. R. (eds.) The Origins of Angiosperms and their Biological Consequences Cambridge University Press, pp. 225–258. ISBN 0-521-32357-6.
- ↑ Lull, R. S. (1908). "The cranial musculature and the origin of the frill in the ceratopsian dinosaurs". American Journal of Science. 4 (25): 387–399. doi:10.2475/ajs.s4-25.149.387.
- ↑ 41,0 41,1 Forster, C. A. (1990). The cranial morphology and systematics of Triceratops, with a preliminary analysis of ceratopsian phylogeny. Ph.D. Dissertation. University of Pennsylvania, Philadelphia. 227 pp.
- ↑ Wheeler, P.E. (1978). "Elaborate CNS cooling structures in large dinosaurs". Nature. 275 (5679): 441–443. doi:10.1038/275441a0. PMID 692723.
- ↑ Davitashvili, L. Sh. (1961). Teoriya Polovogo Otbora (Theory of Sexual Selection). Izdatel'stvo Akademii nauk SSSR. p. 538.
- ↑ Farlow, J.O. and Dodson, P. (1975). "The behavioral significance of frill and horn morphology in ceratopsian dinosaurs". Evolution. 29 (2): 353. doi:10.2307/2407222. JSTOR 2407222.
{{cite journal}}: AS1-onderhoud: meer as een naam (link)
Eksterne skakels[wysig | wysig bron]
- Triceratops-uitstalling Geargiveer 30 September 2013 op Wayback Machine by die Cleveland-museum van Natuurgeskiedenis
- LiveScience: "Facts about Triceratops" by LiveScience.com
- Triceratops, AnimalPlanet
- Smithsonian-uitstalling Geargiveer 2 Februarie 2003 op Wayback Machine
- Triceratops by Internet Archive
- Triceratops in die Dino Directory
 Wikimedia Commons het meer media in die kategorie Triceratops.
Wikimedia Commons het meer media in die kategorie Triceratops.
 Wikispecies het meer inligting verwant aan Triceratops
Wikispecies het meer inligting verwant aan Triceratops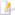 Hierdie artikel is vertaal uit die Engelse Wikipedia
Hierdie artikel is vertaal uit die Engelse Wikipedia