
Kryt-Paleogeen-uitwissing: Verskil tussen weergawes
sjabloon |
Uitgebrei |
||
| Lyn 1: | Lyn 1: | ||
[[Lêer:Impact event.jpg|thumb|’n Kunstenaarsvoorstelling van ’n groot [[Meteoor|meteoriet]] wat die Aarde tref.]] |
[[Lêer:Impact event.jpg|thumb|’n Kunstenaarsvoorstelling van ’n groot [[Meteoor|meteoriet]] wat die Aarde tref.]] |
||
Die '''Kryt-Paleogeen-uitwissing''' (K-Pg-uitwissing) was een van die "Groot Vyf" uitwissingsvoorvalle in die [[Aarde]] se geskiedenis |
Die '''Kryt-Paleogeen-uitwissing''' (K-Pg-uitwissing) was een van die "Groot Vyf" uitwissingsvoorvalle in die [[Aarde]] se geskiedenis toe driekwart van die Aarde se [[plant]]e en [[dier]]e, insluitende alle nie-vlieënde [[dinosourus]]se, 66 miljoen jaar gelede binne ’n geologies kort periode uitgesterf het.<ref name="Renne2013">{{cite journal|last=Renne|first=Paul R.|coauthors=Deino, Alan L.; Hilgen, Frederik J.; Kuiper, Klaudia F.; Mark, Darren F.; Mitchell III, William S.; Morgan, Leah E.; Mundil, Roland; Smit, Jan|title=Time Scales of Critical Events Around the Cretaceous-Paleogene Boundary|journal=Science|date=7 Februarie 2013|volume=339|issue=6120|pages=684–687|doi=10.1126/science.1230492|url=http://www.sciencemag.org/content/339/6120/684|bibcode = 2013Sci...339..684R }}</ref><ref name="Fortey">{{cite book|last=Fortey|first=R|title=Life: A Natural History of the First Four Billion Years of Life on Earth|publisher=Vintage|year=1999|pages=238–260|isbn=978-0-375-70261-7}}</ref> Die uitwissing dui die grens aan tussen die [[Periode (geologie)|geologiese periodes]] [[Kryt (geologie)|Kryt]] (die laaste periode van die [[Mesosoïkum|Mesosoïese Era]]) en [[Paleogeen]] (die eerste periode van die [[Kainosoïkum|Kainosoïese Era]]). |
||
{{Beeld-inligting-uitwissing|float=left}} |
{{Beeld-inligting-uitwissing|float=left}} |
||
| Lyn 25: | Lyn 25: | ||
Spesies wat van fotosintese afhanklik is, het agteruitgegaan of uitgesterf weens stof en as wat die sonlig geblokkeer het. Dit het ’n groot herskommeling teweeggebring in die dominante plantgroepe.<ref name="autogenerated347">{{cite journal|author=Wilf P, Johnson KR |title=Land plant extinction at the end of the Cretaceous: a quantitative analysis of the North Dakota megafloral record |journal=Paleobiology |year=2004 |volume=30 |issue=3 |pages=347–368 |doi = 10.1666/0094-8373(2004)030<0347:LPEATE>2.0.CO;2|issn=0094-8373}}</ref> Organismes wat fotosinteer, het net soos vandag in die laat-Kryt die basis van die voedselketting gevorm. Plantetende diere het uitgesterf toe die plante waarvan hulle afhanklik was, skaars geraak het. Dit het weer tot gevolg gehad dat roofdiere soos ''[[Tyrannosaurus rex]]'' ook uitgesterf het. |
Spesies wat van fotosintese afhanklik is, het agteruitgegaan of uitgesterf weens stof en as wat die sonlig geblokkeer het. Dit het ’n groot herskommeling teweeggebring in die dominante plantgroepe.<ref name="autogenerated347">{{cite journal|author=Wilf P, Johnson KR |title=Land plant extinction at the end of the Cretaceous: a quantitative analysis of the North Dakota megafloral record |journal=Paleobiology |year=2004 |volume=30 |issue=3 |pages=347–368 |doi = 10.1666/0094-8373(2004)030<0347:LPEATE>2.0.CO;2|issn=0094-8373}}</ref> Organismes wat fotosinteer, het net soos vandag in die laat-Kryt die basis van die voedselketting gevorm. Plantetende diere het uitgesterf toe die plante waarvan hulle afhanklik was, skaars geraak het. Dit het weer tot gevolg gehad dat roofdiere soos ''[[Tyrannosaurus rex]]'' ook uitgesterf het. |
||
===Mikrobiota=== |
|||
Die K-Pg-grens verteenwoordig een van die mees drastiese skommelings in die fossielrekord vir verskeie kalkagtige [[Plankton|nanoplankton]], wat die deposito's gevorm het wat aan die Kryt-periode sy naam gee. Dit kan duidelik op spesievlak gesien word. <ref>{{cite journal |author=Pospichal JJ |title=Calcareous nannofossils and clastic sediments at the Cretaceous–Tertiary boundary, northeastern Mexico |year=1996 |journal=Geology |volume=24 |issue=3 |pages=255–258 |url=http://geology.geoscienceworld.org/cgi/content/abstract/24/3/255 |doi=10.1130/0091-7613(1996)024<0255:CNACSA>2.3.CO;2|bibcode = 1996Geo....24..255P }}</ref><ref>{{cite journal |author=Bown P |year=2005 |title=Selective calcareous nannoplankton survivorship at the Cretaceous–Tertiary boundary |journal=Geology |volume=33 |issue=8 |pages=653–656 |url=http://geology.geoscienceworld.org/cgi/content/abstract/33/8/653 | doi =10.1130/G21566.1 |bibcode = 2005Geo....33..653B }}</ref> Die K-Pg-grensrekord van [[sweepdiertjie]]s word nie ewe goed verstaan nie, hoofsaaklik omdat net mikrobiese [[sist]]e ’n fossielrekord het en nie alle sweepdiertjie-spesies sistvormende stadiums het nie; daarom word hul diversiteit waarskynlik onderskat.<ref name="MacLeod">{{cite journal |author=MacLeod N, Rawson PF, Forey PL, Banner FT, Boudagher-Fadel MK, Bown PR, Burnett JA, Chambers, P, Culver S, Evans SE, Jeffery C, Kaminski MA, Lord AR, Milner AC, Milner AR, Morris N, Owen E, Rosen BR, Smith AB, Taylor PD, Urquhart E, Young JR |title=The Cretaceous–Tertiary biotic transition |year=1997 |journal=Journal of the Geological Society |volume=154 |issue=2 |pages=265–292 |url=http://jgs.lyellcollection.org/content/154/2/265.short |doi=10.1144/gsjgs.154.2.0265}}</ref> Onlangse studies het getoon daar was nie groot veranderings onder sweepdiertjies naby die grens nie.<ref>{{cite journal |author=Gedl P |title=Dinoflagellate cyst record of the deep-sea Cretaceous-Tertiary boundary at Uzgru, Carpathian Mountains, Czech Republic |journal=Geological Society, London, Special Publications |year=2004 |volume=230 |pages=257–273 |doi=10.1144/GSL.SP.2004.230.01.13 |bibcode = 2004GSLSP.230..257G }}</ref> |
|||
[[Straaldiertjie]]s het ’n geologiese rekord gelaat sedert die tyd van die [[Ordovisium]], en hul minerale fossiele kom oor die K-Pg-grens voor. Daar is geen bewyse van ’n massauitwissing nie, maar wel van hoë produktiwiteit tydens die vroeë Paleoseen danksy koeler temperature.<ref name="MacLeod"/> Sowat 46% van kristalwier-spesies het die oorgang na die Paleoseen oorleef.<ref name="MacLeod"/><ref>{{cite journal |author=MacLeod N |title=Impacts and marine invertebrate extinctions |year=1998 |journal=Geological Society, London, Special Publications |volume=140 |pages=217–246 |url=http://sp.lyellcollection.org/cgi/content/abstract/140/1/217 | doi = 10.1144/GSL.SP.1998.140.01.16 |issue=1|bibcode = 1998GSLSP.140..217M }}</ref> |
|||
Die voorkoms van plankton[-[foraminifera|foraminifere]] oor die K-Pg-grens word sedert die 1930's bestudeer.<ref>{{cite book | last=Courtillot|first= V | title =Evolutionary Catastrophes: The Science of Mass Extinction | publisher =Cambridge University Press | year=1999 | location =| page =2| url =http://books.google.com/?id=qiegJbafYkUC&pg=PA23&lpg=PA23&dq=foraminifera+kt+1930s | doi =| id = | isbn =0-521-58392-6}}</ref><ref>{{cite journal |author=Glaessner MF |title=Studien über Foraminiferen aus der Kreide und dem Tertiär des Kaukasus: die Foraminiferen der ältesten Tertiärschichten des Nordwestkaukasus |language=German |journal=Проблемс оф палеонтологий - Problems of Paleontology |volume=2–3 |pages=349–410 |year=1937 |oclc=776158910 }}</ref> Talle spesies seebodem-foraminifere is uitgewis, moontlik omdat hulle van organiese afval afhanklik was vir voedingstowwe en die [[biomassa]] in die oseane na verwagting afgeneem het. Aangesien die mariene mikrobiota herstel het, word egter geglo dit het gelei tot ’n toename in die spesies van seebodem-foraminifere.<ref name="MacLeod"/> Die herstel van seebodem-foraminifere het oor verskeie fases gebeur wat verskeie honderdduisende jare geduur het, tot vroeg in die Paleoseen.<ref>{{cite journal |title=Deep-sea benthic foraminiferal recolonisation following a volcaniclastic event in the lower Campanian of the Scaglia Rossa Formation (Umbria-Marche Basin, central Italy)|url=http://www.es.ucl.ac.uk/people/m-kaminski/reprints-pdfs/Galeotti_etal2002.pdf |format=PDF|author=Galeotti S, Bellagamba M, Kaminski MA, and Montanari A |year=2002 |journal=Marine Micropaleontology |volume=44 |pages=57–76 |accessdate=2007-08-19}}</ref><ref>{{cite journal |title=8. Cretaceous to Paleogene benthic foraminifers from the Iberia abyssal plain |author=Kuhnt W, Collins ES |year=1996 |journal=Proceedings of the Ocean Drilling Program, Scientific Results |volume=149 |pages=203–216 |url=http://www-odp.tamu.edu/publications/149_SR/VOLUME/CHAPTERS/SR149_08.PDF |format=PDF|accessdate=2007-08-19}}</ref> |
|||
===Mariene ongewerweldes=== |
|||
[[Lêer:DiscoscaphitesirisCretaceous.jpg|thumb|Die ammoniet ''Discoscaphites iris'', ’n fossiel uit Ripley, [[Mississippi]].]] |
|||
Die gebrek aan fossielrekords van mariene ongewerweldes maak dit moeilik om die omvang van hul uitwissing te bepaal.<ref name="MacLeod"/> |
|||
Sowat 60% van [[Scleractinia]]-koraalgenera uit die laat-Kryt het nie die uitwissingsvoorval oorleef nie. Navorsing toon sowat 98% van koloniale spesies, dié wat in die warm, vlak tropiese water voorgekom het en waarvoor fotosintese belangrik was, <ref>{{cite journal |author=Vescsei A, Moussavian E |title=Paleocene reefs on the Maiella Platform Margin, Italy: An example of the effects of the cretaceous/tertiary boundary events on reefs and carbonate platforms |year=1997 |journal=Facies |volume=36 |issue=1 |pages=123–139 |doi=10.1007/BF02536880}}</ref><ref>{{cite journal |author=Rosen BR, Turnšek D |year=1989 |contribution=Extinction patterns and biogeography of scleractinian corals across the Cretaceous/Tertiary boundary |title=Proceedings of the Fifth International Symposium on Fossil Cnidaria including Archaeocyatha and Spongiomorphs |journal=Memoir of the Association of Australasian Paleontology |location=Brisbane, Queensland |issue=8 |editor1=Jell A |editor2=Pickett JW |pages=355–370}}</ref> is uitgewis. Alleenstaande koraal, wat gewoonlik nie riwwe vorm nie en in dieper, koeler water voorkom, is minder geraak. |
|||
Die getalle genera van [[koppotige]]s, [[stekelhuidige]]s en [[tweekleppige skaaldier]]e het ’n groot afname getoon ná die K-Pg-grens.<ref name="MacLeod"/> Die meeste spesies [[armpotige]]s, ’n klein [[filum]] mariene ongewerweldes, het die uitwissing oorleef en in die vroeë Paleoseen gediversifiseer. |
|||
[[Lêer:RudistCretaceousUAE.jpg|thumb|left|Rudiste, tweekleppige weekdiere, uit die laat-Kryt in die [[Verenigde Arabiese Emirate]].]] |
|||
Buiten [[nautiloïdee|nautiloïdeë]] (wat verteenwoordig word deur die moderne orde [[Nautilida]]) en [[inkvis]]se (wat reeds gesplits het in moderne [[seekat]]te, [[pylinkvis]]se en [[tjokka]]) het alle ander spesies van die [[weekdier]]klas [[Cephalopoda]] by die K-Pg-grens uitgesterf. Dit sluit in die ekologies belangrike [[Belemnoidea]], sowel as die [[Ammonoidea]] of ammoniete, ’n subklas baie uiteenlopende, talryke en wydverspreide hardedop-koppotiges. Navorsers wys daarop dat die voortplantingstrategie van die oorlewende Nautilida (minder en groter eiers) ’n rol gespeel het in die feit dat hulle oorleef het en die Ammonoidea nie. Laasgenoemde het meer en kleiner eiers gelê, en die uitwissingsvoorval het ’n verwoestende uitwerking daarop gehad. Ná die uitwissing van die Ammonoidea het die Nautilida ’n evolusie ondergaan met dopvorms en kompleksiteite wat voorheen onbekend was in dié subklas.<ref>{{cite journal |author=Ward PD, Kennedy WJ, MacLeod KG, Mount JF |year=1991 |title=Ammonite and inoceramid bivalve extinction patterns in Cretaceous/Tertiary boundary sections of the Biscay region (southwestern France, northern Spain) |journal=Geology |volume=19 |issue=12 |pages=1181–1184 |url=http://geology.geoscienceworld.org/cgi/content/abstract/19/12/1181 |doi=10.1130/0091-7613(1991)019<1181:AAIBEP>2.3.CO;2|bibcode = 1991Geo....19.1181W }}</ref><ref>{{cite journal |author=Harries PJ, Johnson KR, Cobban WA, Nichols DJ |year=2002 |title=Marine Cretaceous-Tertiary boundary section in southwestern South Dakota: Comment and Reply |journal=Geology |volume=30 |issue=10 |pages=954–955 |doi=10.1130/0091-7613(2002)030<0955:MCTBSI>2.0.CO;2 |issn=0091-7613|bibcode = 2002Geo....30..954H }}</ref> |
|||
Sowat 35% van die genera stekelhuidiges het uitgesterf by die K-Pg-grens, hoewel taksons wat by lae breedteliggings in vlak water floreer het, die swaarste getref is.<ref>{{cite journal |author=Vescsei A, Moussavian E |title=Paleocene reefs on the Maiella Platform Margin, Italy: An example of the effects of the cretaceous/tertiary boundary events on reefs and carbonate platforms |year=1997 |journal=Facies |volume=36 |issue=1 |pages=123–139 |doi=10.1007/BF02536880}}</ref><ref>{{cite journal |author=Rosen BR, Turnšek D |year=1989 |contribution=Extinction patterns and biogeography of scleractinian corals across the Cretaceous/Tertiary boundary |title=Proceedings of the Fifth International Symposium on Fossil Cnidaria including Archaeocyatha and Spongiomorphs |journal=Memoir of the Association of Australasian Paleontology |location=Brisbane, Queensland |issue=8 |editor1=Jell A |editor2=Pickett JW |pages=355–370}}</ref> Diepwater-stekelhuidiges by medium breedteliggings is baie minder geraak. |
|||
===Visse=== |
|||
Daar is heelwat fossielrekords van [[kaakdier]]visse oor die K-Pg-grens, wat ’n goeie aanduiding van uitwissingspatrone van dié klasse mariene [[gewerwelde]]s gee. In die klas Chondrichthyes het sowat 80% van die [[haai]]-, [[kraakbeenvis]]- en [[Rajidae]]-families die uitwissing oorleef,<ref name="MacLeod"/> en meer as 90% van die [[Teleostei]] (beenvis)-families.<ref>{{cite book |last=Patterson|first=C |year=1993 |title=Osteichthyes: Teleostei. In: The Fossil Record 2 (Benton, MJ, editor) |publisher=Springer |pages=621–656 |isbn=0-412-39380-8}}</ref> |
|||
===Ongewerweldes op land=== |
|||
[[Insek]]skade aan gefossileerde blare van blomplante op 14 terreine in Noord-Amerika is ontleed om ’n idee te kry van die diversiteit van insekte oor die K-Pg-grens heen. Navorsers het gevind Kryt-terreine het voor die uitwissing ’n groot verskeidenheid plante gehad met heelwat tekens van insekte wat hulle vreet. Tydens die vroeë Paleoseen was daar egter min tekens van plundering deur insekte, selfs 1,7 miljoen jaar ná die uitwissing.<ref name="Labandieraetal">{{cite journal| author =Labandeira Conrad C, Johnson Kirk R, Wilf Peter | title =Impact of the terminal Cretaceous event on plant–insect associations| journal =Proceedings of the National Academy of Sciences of the United States of America| volume =99| issue =4| pages =2061–2066| year =2002| url =http://www.geosc.psu.edu/~pwilf/LabandeiraJohnsonWilf%202002PNAS.pdf| format=PDF |
|||
| doi = 10.1073/pnas.042492999 | pmid =11854501| pmc =122319 |bibcode = 2002PNAS...99.2061L }}</ref><ref>{{cite journal |title=Decoupled Plant and Insect Diversity After the End-Cretaceous Extinction |author=Wilf P, Labandeira CC, Johnson KR, Ellis B |year=2006 |journal=Science |volume=313 |issue=5790 |pages=1112–1115 |doi=10.1126/science.1129569 |pmid=16931760|bibcode = 2006Sci...313.1112W }}</ref> |
|||
===Landplante=== |
|||
Daar is oorweldigende bewyse van ’n wêreldwye ontwrigting van plantgroepe by die K-Pg-grens.<ref name="Nichols, D. J 2008"/><ref name="Nichols, D. J 2008"/><ref name="Vajdaetal">{{cite journal| author =Vajda Vivi, Raine J Ian, Hollis Christopher J | title =Indication of Global Deforestation at the Cretaceous–Tertiary Boundary by New Zealand Fern Spike | journal =Science | volume =294 | issue =5547| pages =1700–1702| year =2001| url =http://www.sciencemag.org/cgi/content/abstract/294/5547/1700 | doi =10.1126/science.1064706 | pmid =11721051|bibcode = 2001Sci...294.1700V }}</ref><ref>{{cite journal | author = Wilf P., Johnson K. R. | year = 2004 | title = Land plant extinction at the end of the Cretaceous: a quantitative analysis of the North Dakota megafloral record | url = | journal = Paleobiology | volume = 30 | issue = 3| pages = 347–368 }}</ref> Uitwissings blyk duidelik uit die bestudering van fossiele stuifmeel en blare.<ref name="Nichols, D. J 2008"/> In Noord-Amerika dui die data op ’n enorme verwoesting en massauitwissing by die K-Pg-grens, maar daar was al voor die grens aansienlike veranderings in die plantegroei.<ref name="Nichols, D. J 2008"/><ref>{{cite book |last=Johnson|first= KR|coauthors=Hickey LJ |year=1991 |title=Megafloral change across the Cretaceous Tertiary boundary in the northern Great Plains and Rocky Mountains. In: Global Catastrophes in Earth History: An Interdisciplinary Conference on Impacts, Volcanism, and Mass Mortality, Sharpton VI and Ward PD (redakteurs) |publisher=Geological Society of America |isbn=978-0-8137-2247-4}}</ref> Sowat 57% van plantspesies op dié vasteland het uitgesterf. In die verre suide soos in [[Nieu-Seeland]] en op [[Antarktika]], het die massauitsterwing van plante nie ’n beduidende omwenteling in spesies teweeggebring nie, maar wel drastiese en korttermynveranderinge in die relatiewe oorvloed van plantgroepe.<ref name="Labandieraetal"/><ref>{{cite book |last=Askin|first= RA|coauthors=Jacobson SR |year=1996 |title=Palynological change across the Cretaceous–Tertiary boundary on Seymour Island, Antarctica: environmental and depositional factors. In: Cretaceous–Tertiary Mass Extinctions: Biotic and Environmental Changes, Keller G, MacLeod N (editors) |publisher=WW Norton |isbn=978-0-393-96657-2}}</ref> In sommige streke het die herstel van plante begin met die terugkeer van [[varing]]spesies.<ref name="SchultzDhondt">{{cite journal | author =Schultz PH, D'Hondt S| title =Cretaceous–Tertiary (Chicxulub) impact angle and its consequences | journal =Geology | volume =24 | issue =11 | pages =963–967 | year=1996 | url=http://geology.geoscienceworld.org/cgi/content/abstract/24/11/963 | doi =10.1130/0091-7613(1996)024<0963:CTCIAA>2.3.CO;2| id =| accessdate =2007-07-05|bibcode = 1996Geo....24..963S }}</ref> Die patrone wissel egter – verskillende varingspesies was in verskillende gebiede verantwoordelik vir die oplewing en is sekere gebiede is geen oplewing te bespeur nie. |
|||
Vanweë die grootskaalse uitwissing van plante was daar ’n groot toename in [[saprofyt]]-organismes soos [[fungus]]se wat nie fotosintese nodig het nie en hul voedingstowwe uit verrottende plante kry. Die oorheersing van fungusse het net ’n paar jaar geduur terwyl daar baie organiese matriaal was wat voeding verskaf het. Toe die atmosfeer weer skoon was, het fotosintetiese plante soos varings en ander soorte teruggekeer.<ref>{{cite journal |author=Vajda V, McLoughlin S |year=2004 |title=Fungal Proliferation at the Cretaceous–Tertiary Boundary |journal=Science |url=http://www.geol.lu.se/personal/VIV/Vajda%20&McLoughlin%202004.pdf |format=PDF |volume=303 |pages=1489–1490 |doi=10.1126/science.1093807 |accessdate=2007-07-07 |pmid=15001770 |issue=5663}}</ref> |
|||
[[Polyploïdie]] (die verskynsel dat nakomelinge ’n groter getal [[chromosoom|chromosome]] as die ouers het) het skynbaar die vermoë van blomplante verhoog om die uitwissing te oorleef, moontlik aangesien die bykomende kopieë van die [[genoom]] wat sulke plante besit, hulle in staat gestel het om makliker by die omgewingsveranderings aan te pas. <ref name='Fawcett2009'>Fawcett, J. A.; Maere, S.; Van de Peer, Y. (April 2009). "Plants with double genomes might have had a better chance to survive the Cretaceous–Tertiary extinction event". Proceedings of the National Academy of Sciences of the United States of America 106 (14): 5737–5742.Bibcode:2009PNAS..106.5737F. doi:10.1073/pnas.0900906106.ISSN 0027-8424. PMC 2667025</ref> |
|||
===Amfibieë=== |
|||
Daar is min getuienis van die uitwissing van amfibieë by die K-Pg-grens. Volgens ’n ondersoek in Montana van fossiele gewerweldes oor die K-Pg-grens heen is gevind dat geen amfibiese spesies uitgewis is nie.<ref name="Archibald">{{cite journal |author=Archibald JD, Bryant LJ |year=1990 |title=Differential Cretaceous–Tertiary extinction of nonmarine vertebrates; evidence from northeastern Montana. In: Global Catastrophes in Earth History: an Interdisciplinary Conference on Impacts, Volcanism, and Mass Mortality (Sharpton VL and Ward PD, editors) |journal=Geological Society of America, Special Paper |volume=247 |pages=549–562}}</ref> Daar is egter soorte amfibieë wat nie deel van die ondersoek was nie wat onbekend was in die Paleoseen, soos die padda ''Theatonius lancensis'';<ref>Estes, R. (1964). "Fossil vertebrates from the Late Cretaceous Lance Formation, Eastern Wyoming." University of California Publications, Department of Geological Sciences 49: 1-180.</ref> en die salamander ''Albanerpeton galaktion'';<ref>{{cite journal | author = Gardner J. D. | year = 2000 | title = Albanerpetontid amphibians from the Upper Cretaceous (Campanian and Maastrichtian) of North America | url = | journal = Geodiversitas | volume = 22 | issue = 3| pages = 349–388 }}</ref> dus lyk dit of sommige amfibieë wel uitgesterf het. |
|||
===Nie-archosourus-reptiele=== |
|||
Die twee nie-archosourus-reptiele, [[Testudines]] ([[skilpad|skilpaaie]]) en [[Lepidosauria]] ([[slange]]e, [[akkedis]]se en [[wurmakkedis]]se), asook [[Choristodera]] (semi-waterdiere wat in die vroeë [[Mioseen]] uitgesterf het), het die uitwissing oorleef.<ref name="MacLeod"/> Meer as 80% van die Kryt se skilpaaie het oorleef. Daarbenewens het al ses skilpadfamilies wat aan die einde van die Kryt voorgekom het, oorleef tot in die Paleogeen en hulle word deur huidige spesies verteenwoordig.<ref name="Novacek">{{cite journal |title=100 Million Years of Land Vertebrate Evolution: The Cretaceous-Early Tertiary Transition |author=Novacek MJ |journal=Annals of the Missouri Botanical Garden |volume=86 |issue=2 |year=1999 |pages=230–258 |doi=10.2307/2666178 |jstor=2666178}}</ref> |
|||
Die orde [[Squamata]], wat vandag verteenwoordig word deur akkedisse, slange en wurmakkedisse, het verskeie ekologiese nisse gevul tydens die [[Jura]] en was suksesvol tydens die Kryt. Hulle het die uitwissing oorleef en is tans die suksesvolste en uiteenlopendse groep lewende reptiele met meer as 6 000 spesies. Nie ’n enkele familie van aardlewende Squamata is uitgewis nie en fossiele getuienis dui daarop dat hulle geen noemenswaardige afname in getalle beleef het nie. Hul klein grootte, aanpasbare metabolisme en vermoë om na gunstiger habitats te trek, was sleutelfaktore in die feit dat hulle oorleef het.<ref name="MacLeod"/><ref name="Novacek"/> Reusagtige nie-Archosourus-waterreptiele soos [[Mosasauridae]] en [[Plesiosauria]], wat die belangrikste mariene roofdiere van hul tyd was, het aan die einde van die Kryt uitgesterf.<ref name="ChatterjeeSmall">{{cite journal | author=Chatterjee S, Small BJ | title =New plesiosaurs from the Upper Cretaceous of Antarctica| journal =Geological Society, London, Special Publications| volume =47| issue =1| pages =197–215| year =1989| url =http://sp.lyellcollection.org/cgi/content/abstract/47/1/197 |
|||
| doi = 10.1144/GSL.SP.1989.047.01.15 | accessdate =2007-07-04|bibcode = 1989GSLSP..47..197C }}</ref><ref>{{cite journal |author=O'Keefe FR |year=2001 |title=A cladistic analysis and taxonomic revision of the Plesiosauria (Reptilia: Sauropterygia) |journal=Acta Zoologica Fennica |volume=213 |pages=1–63}}</ref> Hulle was waarskynlik minder in staat om by veranderende toestande aan te pas weens hul grootte. |
|||
===Archosourusse=== |
|||
Die archosourus-klade sluit twee huidige ordes in, [[Crocodilia]] (waarvan [[Alligatoridae]], [[Crocodylidae]] en [[Gavialidae]] die enigste oorlewende families is) en dinosourusse (waarvan voëls die enigste oorlewende lede is), asook die uitgestorwe nie-vlieënde dinosourusse en die pterosourus. |
|||
====Voëls==== |
|||
Die meeste paleontoloë beskou voëls as die enigste oorlewende dinosourusse. Alle nie-[[Neornithes]]-voëls het egter uitgesterf, insluitende florerende groepe soos [[Enantiornithes]] en [[Hesperornithes]].<ref>{{cite journal |author=Hou L, Martin M, Zhou Z, Feduccia A |year=1996 |title=Early Adaptive Radiation of Birds: Evidence from Fossils from Northeastern China |journal=Science |volume=274 |issue=5290 |pages=1164–1167 |doi=10.1126/science.274.5290.1164 |pmid=8895459 |bibcode = 1996Sci...274.1164H }}</ref> Verskeie ondersoeke van voëlfossiele toon die splitsing van spesies voor die K-Pg-grens en dat verwante van [[eend]]e, [[hoender]]s en [[loopvoël]]s saam met nie-vlieënde dinosorusse bestaan het.<ref>{{cite journal |title=Definitive fossil evidence for the extant avian radiation in the Cretaceous |author=Clarke JA, Tambussi CP, Noriega JI, Erickson GM, Ketcham RA |year=2005 |journal=Nature |volume=433 |issue=7023 |pages=305–308 |doi=10.1038/nature03150 |pmid=15662422 |bibcode = 2005Natur.433..305C }}</ref> Groot versamelings voëlfossiele lewer voldoende bewys dat daar in die Kryt ’n groot verskeidenheid spesies was wat nie die Paleogeen gehaal het nie. ’n Baie klein deeltjie van Kryt se die voëls het dus oorleef en uit hulle het vandag se voëls ontwikkel.<ref name=longrichetal2011>{{cite journal | doi = 10.1073/pnas.1110395108 | last1 = Longrich | first1 = Nicholas R. | last2 = Tokaryk | first2 = Tim | last3 = Field | first3 = Daniel J. | year = 2011 | title = Mass extinction of birds at the Cretaceous–Paleogene (K–Pg) boundary. | url = http://www.pnas.org/content/108/37/15253 | journal = Proceedings of the National Academy of Sciences | volume = 108 | issue = 37 | pages = 15253–15257|bibcode = 2011PNAS..10815253L }}</ref><ref>{{cite web | url = http://www.sciencedaily.com/releases/2011/09/110919151315.htm | title = Primitive Birds Shared Dinosaurs' Fate | date = 20 September 2011 | publisher = Science Daily | accessdate= 20 September 2011}}</ref> Die enigste subklas voëls wat beslis die uitwissing oorleef het, is [[Neornithes]] (hoewel een Paleogeen-spesie, ''[[Qinornis paleocenica]]'', verteenwoordigend kan wees van ’n nie-Neornithes-voël wat oorleef het.<ref name=longrichetal2011/> Neornithes kon vermoedelik die uitwissing oorleef danksy hul vermoë om te kan duik, swem of skuiling soek in water en moeraslande. Baie spesies kan ook neste bou en in gate in bome of mierneste bly, wat hulle ook kon beskerm het. Hulle het ná die uitwissing ekologiese nisse gevul wat leeg gelaat is deur die uitsterwing van die nie-vlieënde dinosourusse.<ref name="Robertson">{{cite journal |title= Survival in the first hours of the Cenozoic |author=Robertson DS, McKenna MC, Toon OB, Hope S, Lillegraven JA |journal=GSA Bulletin |year=2004 |volume=116 |issue=5–6 |pages=760–768 |doi=10.1130/B25402.1 |url=http://www.ugcs.caltech.edu/~presto/cenozoic.pdf |format=PDF|accessdate=2007-08-31|bibcode = 2004GSAB..116..760R }}</ref> |
|||
====Nie-vlieënde dinosourusse==== |
|||
[[Lêer:Tyrannosaurus Rex Holotype.jpg|thumb|''Tyrannosaurus'' was een van die laaste dinosourusse op Aarde voor die uitwissing.]] |
|||
Buiten ’n paar omstrede bewerings, stem wetenskaplikes saam dat alle nie-vlieënde dinosourusse uitgesterf het by die K-Pg-grens. Die dinisourus-fossielrekord is geïnterpreteer as dat daar óf ’n afname óf geen afname in verskeidenheid was in die laaste paar miljoen jaar van die Kryt, en dit kan wees dat die gehalte van die fossielrekord net te swak was om tussen die twee opsies te onderskei.<ref name=AF04>{{cite book |first=Archibald|last=David|coauthors=David Fastovsky|editor=Weishampel David B, Dodson Peter, Osmólska Halszka (eds.) |title=The Dinosauria |edition=2nd |year=2004 |publisher=University of California Press |location=Berkeley |isbn=0-520-24209-2 |pages=672–684 |chapter=Dinosaur Extinction |chapterurl=http://www.bio.sdsu.edu/faculty/archibald/ArchFast.pdf }}</ref> Aangesien daar geen bewys is dat nie-vlieënde dinosourusse gate kon grawe, swem of duik nie, was hulle nie in staat om hulself te beskerm teen die omgewingstres wat by die K-Pg-grens voorgekom het nie. Kleiner diere (benewens voëls) kon dalk oorleef het, maar hulle sou geen kos gehad het nie aangesien plante skaars sou gewees het vir plantvretende diere en dit sou weer ’n voedseltekort veroorsaak het vir vleisvretende diere.<ref name="Robertson"/> |
|||
Die toenemende konsensus oor die [[endotermie]] ("warmbloedigheid") van dinosourusse help om hul uitwissing te verstaan, terwyl hul verwante die krokodille oorleef het. Die [[ektotermie]]se ("koudbloedige") krokodille sou ’n klein behoefte aan kos gehad het aangesien hulle verskeie maande sonder kos kan klaarkom, terwyl endotermiese diere van dieselfde grootte meer kos nodig sou gehad het vir hul vinniger metabolisme. Nie-vlieënde dinosourusse sou dus uitgesterf het weens ’n tekort aan kos,<ref name="autogenerated347"/> terwyl sommige krokodille oorleef het. Die oorlewing van ander warmbloedige diere, soos soogdiere, kon danksy ander redes gewees het soos hul kleiner behoefte aan kos vanweë hul kleiner grootte in dié tyd.<ref name="springerlink.com">{{cite book |last=Ocampo|first=A|coauthors=Vajda V, Buffetaut E |chapter=Unravelling the Cretaceous–Paleogene (K–T) turnover, evidence from flora, fauna and geology in biological processes associated with impact events |editor=Cockell C, Gilmour I, Koeberl C |title=Biological Processes Associated with Impact Events |publisher=SpringerLink |year=2006 |pages=197–219 |isbn=978-3-540-25735-6|url=http://www.springerlink.com/content/vw75014157p2p278/}}</ref> |
|||
===Soogdiere=== |
|||
Alle belangrike soogdierlyne uit die Kryt, insluitende [[kloaakdier]]e (eierlêende soogdiere), [[Multituberculata]], [[buideldier|buidel-]] en [[plasentadier]]e, [[Dryolestoidea]]<ref>{{cite journal|author=Gelfo JN and Pascual R|year=2001|title=''Peligrotherium tropicalis'' (Mammalia, Dryolestida) from the early Paleocene of Patagonia, a survival from a Mesozoic Gondwanan radiation|journal=Geodiversitas|volume=23|pages=369–379|url=http://www.mnhn.fr/publication/geodiv/g01n3a4.pdf }}</ref> en [[Gondwanatheria]]<ref>{{cite journal|author=Goin FJ, Reguero MA, Pascual R, von Koenigswald W, Woodburne MO, Case JA, Marenssi SA, Vieytes C, Vizcaíno SF|year=2006|title=First gondwanatherian mammal from Antarctica|journal=Geological Society, London, Special Publications|volume=258|pages=135–144|doi=10.1144/GSL.SP.2006.258.01.10|bibcode = 2006GSLSP.258..135G }}</ref> het die uitwissing oorleef hoewel daar ook groot verliese was. Buideldiere het grootliks uit Noord-Amerika verdwyn, en die Asiatiese [[Deltatheroida]], primitiewe verwante van bestaande buideldiere, is uitgewis.<ref name=classofmamm>{{cite book|last=McKenna|first= MC|coauthors=Bell SK|title=Classification of mammals: above the species level|publisher=Columbia University Press|year=1997|isbn=978-0-231-11012-9}}</ref> |
|||
Die soogdiere wat die uitwissing oorleef het, was oor die algemeen klein – omtrent so groot soos rotte. Dit sou hulle gehelp het om skuiling te vind. Daar word ook gemeen baie van hulle kon gate grawe of was semi-waterdiere omdat daar vandag verskeie sulke soogdierspesies is. Dit sou bykomende beskutting gebied het teen omgewingstres.<ref name="Robertson"/> |
|||
==Chicxulub-asteroïde== |
==Chicxulub-asteroïde== |
||
Wysiging soos op 21:43, 28 Januarie 2014

Die Kryt-Paleogeen-uitwissing (K-Pg-uitwissing) was een van die "Groot Vyf" uitwissingsvoorvalle in die Aarde se geskiedenis toe driekwart van die Aarde se plante en diere, insluitende alle nie-vlieënde dinosourusse, 66 miljoen jaar gelede binne ’n geologies kort periode uitgesterf het.[1][2] Die uitwissing dui die grens aan tussen die geologiese periodes Kryt (die laaste periode van die Mesosoïese Era) en Paleogeen (die eerste periode van die Kainosoïese Era).



In die geologiese rekord word die uitwissing aangedui deur ’n dun rotslaag, die Kryt-Paleogeen-grens, wat regdeur die wêreld in see- en aardrotse gevind word. Dit bevat hoë vlakke van die metaal iridium, wat seldsaam in die aardkors is, maar volop in asteroïdes voorkom.
Daar word algemeen geglo die K-Pg-uitwissing is veroorsaak deur ’n groot komeet of asteroïde wat die Aarde getref het en ’n katastrofale uitwerking op die omgewing gehad het, insluitende ’n lang impakwinter (weens stof en as wat die Son se strale afkeer), wat dit vir plante onmoontlik gemaak het om fotosintese uit te voer.[3] Dié teorie is ondersteun deur die ontdekking van die 18 km wye Chicxulub-krater in die laat 1970's in die Golf van Mexiko,[4] wat onteenseglik bewys het dat die K-Pg-grens afval van ’n asteroïde-impak verteenwoordig.[5] Die feit dat die uitwissing op dieselfde tyd plaasgevind het as die impak, is ook ’n bewys dat die uitwissing deur die asteroïde veroorsaak is.[5]
Daar is egter wetenskaplikes wat glo die uitwissing is veroorsaak of vererger deur ander faktore soos vulkaniese uitbarstings,[6] klimaatsveranderings en/of ’n verandering in seevlakke.
’n Groot verskeidenheid spesies het in die K-Pg-uitwissing omgekom. Die bekendste slagoffers was die nie-vlieënde dinosourusse. Ander organismes is egter ook erg geraak, soos soogdiere, voëls,[7] pterosourusse, akkedisse,[8] insekte,[9][10] en plante.[11] In die oseane is groot seereptiele, visse,[12] haaie, weekdiere en baie soorte plankton swaar getref. Daar word geraam dat tot 75% van die spesies op Aarde kon verdwyn het.[13]
Die verwoesting wat deur die uitwissing veroorsaak is, het egter ook evolusiegeleenthede geskep. Ná die voorval het baie groepe skielike en groot veranderings ondergaan en is nuwe lewensvorme geskep wat die ekologiese nisse gevul het wat deur die uitwissing leeg gelaat is. Soogdiere het veral in die Paleogeen gediversifiseer[14] en nuwe vorms aangeneem soos perde, walvisse, vlermuise en primate. Voëls,[15] visse[16] en dalk akkedisse[8] het ook versprei.
Patrone van uitwissing
Die K-Pg-uitwissing was ’n wêreldwye gebeurtenis. Dit het al die kontinente oënskynlik terselfdertyd getref. Dinosourusse het byvoorbeeld voorgekom van Noord-Amerika tot Europa, Asië, Afrika, Suid-Amerika en Antarktika,[17] maar word sedert die Kainosoïkum nêrens ter wêreld aangetref nie. Net so het stuifmeelfossiele getoon plante in verafgeleë streke soos Nieu-Mexiko, Alaska, Sjina en Nieu-Seeland is geraak.[11] Die uitwissing het ook beide die see en land gelyktydig getref.
Die fossielrekords toon die uitwissing het oor ’n relatief kort tydperk van 1 000 jaar of minder gebeur. In sommige gevalle kan fossiele in K-Pg-rotse op baie fyn skaal bestudeer word – sentimeter vir sentimeter. Fossiele van stuifmeel en plankton toon dat die ekosisteem op plekke relatief stabiel was, tot met die K-Pg-grens, toe baie spesies skielik verdwyn het. Vir groepe met ’n swakker fossielrekord, soos dinosourusse, is dit onwaarskynlik dat fossiele net onder die K-Pg-grens gepreserveer sal word. Daar is byvoorbeeld net ’n paar dosyn Tyrannosourus-geraamtes gevind, en die kans om een te vind ’n paar sentimeter onder die grens, is klein. Hierdie effek, bekend as die Signor-Lipps-effek, veroorsaak dat dit lyk of baie spesies voor die the K-Pg-grens verdwyn het, en dit skep die illusie van geleidelike uitwissing.
Beter monsters het egter gewys dat groepe wat voorheen vermoedelik geleidelik uitgesterf het, soos dinosourusse, eintlik skielik naby die K-Pg-grens verdwyn het. Wanneer erosie fossiele uit ouer deposito's ontbloot en hulle dan vasgevang word in jonger deposito's, kan dit ook lyk of die uitwissing geleidelik gebeur het. So is dinosourus-fossiele in Montana gevind langs fossiele van soogdiere; dit laat dit voorkom of dinosourusse nog voorgekom het terwyl soogdiere versprei het. Sulke fossiele is egter maklik herkenbaar omdat hulle skaars en dikwels beskadig is. Dié patrone is baie belangrik om die redes vir uitwissings te verstaan.
Die uitwissing was ook baie selektief. Sommige groepe is feitlik nie geraak nie, ander is grootliks verwoes en nog ander is heeltemal uitgewis. Baie spesies alligators, skilpaaie en salamanders het byvoorbeeld oorleef. Soogdiere, voëls en akkedisse is erg geraak. Nie-vlieënde dinosourusse en die pterosourusse is heeltemal uitgewis.
Spesies wat van fotosintese afhanklik is, het agteruitgegaan of uitgesterf weens stof en as wat die sonlig geblokkeer het. Dit het ’n groot herskommeling teweeggebring in die dominante plantgroepe.[18] Organismes wat fotosinteer, het net soos vandag in die laat-Kryt die basis van die voedselketting gevorm. Plantetende diere het uitgesterf toe die plante waarvan hulle afhanklik was, skaars geraak het. Dit het weer tot gevolg gehad dat roofdiere soos Tyrannosaurus rex ook uitgesterf het.
Mikrobiota
Die K-Pg-grens verteenwoordig een van die mees drastiese skommelings in die fossielrekord vir verskeie kalkagtige nanoplankton, wat die deposito's gevorm het wat aan die Kryt-periode sy naam gee. Dit kan duidelik op spesievlak gesien word. [19][20] Die K-Pg-grensrekord van sweepdiertjies word nie ewe goed verstaan nie, hoofsaaklik omdat net mikrobiese siste ’n fossielrekord het en nie alle sweepdiertjie-spesies sistvormende stadiums het nie; daarom word hul diversiteit waarskynlik onderskat.[21] Onlangse studies het getoon daar was nie groot veranderings onder sweepdiertjies naby die grens nie.[22]
Straaldiertjies het ’n geologiese rekord gelaat sedert die tyd van die Ordovisium, en hul minerale fossiele kom oor die K-Pg-grens voor. Daar is geen bewyse van ’n massauitwissing nie, maar wel van hoë produktiwiteit tydens die vroeë Paleoseen danksy koeler temperature.[21] Sowat 46% van kristalwier-spesies het die oorgang na die Paleoseen oorleef.[21][23]
Die voorkoms van plankton[-[foraminifera|foraminifere]] oor die K-Pg-grens word sedert die 1930's bestudeer.[24][25] Talle spesies seebodem-foraminifere is uitgewis, moontlik omdat hulle van organiese afval afhanklik was vir voedingstowwe en die biomassa in die oseane na verwagting afgeneem het. Aangesien die mariene mikrobiota herstel het, word egter geglo dit het gelei tot ’n toename in die spesies van seebodem-foraminifere.[21] Die herstel van seebodem-foraminifere het oor verskeie fases gebeur wat verskeie honderdduisende jare geduur het, tot vroeg in die Paleoseen.[26][27]
Mariene ongewerweldes

Die gebrek aan fossielrekords van mariene ongewerweldes maak dit moeilik om die omvang van hul uitwissing te bepaal.[21]
Sowat 60% van Scleractinia-koraalgenera uit die laat-Kryt het nie die uitwissingsvoorval oorleef nie. Navorsing toon sowat 98% van koloniale spesies, dié wat in die warm, vlak tropiese water voorgekom het en waarvoor fotosintese belangrik was, [28][29] is uitgewis. Alleenstaande koraal, wat gewoonlik nie riwwe vorm nie en in dieper, koeler water voorkom, is minder geraak.
Die getalle genera van koppotiges, stekelhuidiges en tweekleppige skaaldiere het ’n groot afname getoon ná die K-Pg-grens.[21] Die meeste spesies armpotiges, ’n klein filum mariene ongewerweldes, het die uitwissing oorleef en in die vroeë Paleoseen gediversifiseer.

Buiten nautiloïdeë (wat verteenwoordig word deur die moderne orde Nautilida) en inkvisse (wat reeds gesplits het in moderne seekatte, pylinkvisse en tjokka) het alle ander spesies van die weekdierklas Cephalopoda by die K-Pg-grens uitgesterf. Dit sluit in die ekologies belangrike Belemnoidea, sowel as die Ammonoidea of ammoniete, ’n subklas baie uiteenlopende, talryke en wydverspreide hardedop-koppotiges. Navorsers wys daarop dat die voortplantingstrategie van die oorlewende Nautilida (minder en groter eiers) ’n rol gespeel het in die feit dat hulle oorleef het en die Ammonoidea nie. Laasgenoemde het meer en kleiner eiers gelê, en die uitwissingsvoorval het ’n verwoestende uitwerking daarop gehad. Ná die uitwissing van die Ammonoidea het die Nautilida ’n evolusie ondergaan met dopvorms en kompleksiteite wat voorheen onbekend was in dié subklas.[30][31]
Sowat 35% van die genera stekelhuidiges het uitgesterf by die K-Pg-grens, hoewel taksons wat by lae breedteliggings in vlak water floreer het, die swaarste getref is.[32][33] Diepwater-stekelhuidiges by medium breedteliggings is baie minder geraak.
Visse
Daar is heelwat fossielrekords van kaakdiervisse oor die K-Pg-grens, wat ’n goeie aanduiding van uitwissingspatrone van dié klasse mariene gewerweldes gee. In die klas Chondrichthyes het sowat 80% van die haai-, kraakbeenvis- en Rajidae-families die uitwissing oorleef,[21] en meer as 90% van die Teleostei (beenvis)-families.[34]
Ongewerweldes op land
Insekskade aan gefossileerde blare van blomplante op 14 terreine in Noord-Amerika is ontleed om ’n idee te kry van die diversiteit van insekte oor die K-Pg-grens heen. Navorsers het gevind Kryt-terreine het voor die uitwissing ’n groot verskeidenheid plante gehad met heelwat tekens van insekte wat hulle vreet. Tydens die vroeë Paleoseen was daar egter min tekens van plundering deur insekte, selfs 1,7 miljoen jaar ná die uitwissing.[35][36]
Landplante
Daar is oorweldigende bewyse van ’n wêreldwye ontwrigting van plantgroepe by die K-Pg-grens.[11][11][37][38] Uitwissings blyk duidelik uit die bestudering van fossiele stuifmeel en blare.[11] In Noord-Amerika dui die data op ’n enorme verwoesting en massauitwissing by die K-Pg-grens, maar daar was al voor die grens aansienlike veranderings in die plantegroei.[11][39] Sowat 57% van plantspesies op dié vasteland het uitgesterf. In die verre suide soos in Nieu-Seeland en op Antarktika, het die massauitsterwing van plante nie ’n beduidende omwenteling in spesies teweeggebring nie, maar wel drastiese en korttermynveranderinge in die relatiewe oorvloed van plantgroepe.[35][40] In sommige streke het die herstel van plante begin met die terugkeer van varingspesies.[41] Die patrone wissel egter – verskillende varingspesies was in verskillende gebiede verantwoordelik vir die oplewing en is sekere gebiede is geen oplewing te bespeur nie.
Vanweë die grootskaalse uitwissing van plante was daar ’n groot toename in saprofyt-organismes soos fungusse wat nie fotosintese nodig het nie en hul voedingstowwe uit verrottende plante kry. Die oorheersing van fungusse het net ’n paar jaar geduur terwyl daar baie organiese matriaal was wat voeding verskaf het. Toe die atmosfeer weer skoon was, het fotosintetiese plante soos varings en ander soorte teruggekeer.[42]
Polyploïdie (die verskynsel dat nakomelinge ’n groter getal chromosome as die ouers het) het skynbaar die vermoë van blomplante verhoog om die uitwissing te oorleef, moontlik aangesien die bykomende kopieë van die genoom wat sulke plante besit, hulle in staat gestel het om makliker by die omgewingsveranderings aan te pas. [43]
Amfibieë
Daar is min getuienis van die uitwissing van amfibieë by die K-Pg-grens. Volgens ’n ondersoek in Montana van fossiele gewerweldes oor die K-Pg-grens heen is gevind dat geen amfibiese spesies uitgewis is nie.[44] Daar is egter soorte amfibieë wat nie deel van die ondersoek was nie wat onbekend was in die Paleoseen, soos die padda Theatonius lancensis;[45] en die salamander Albanerpeton galaktion;[46] dus lyk dit of sommige amfibieë wel uitgesterf het.
Nie-archosourus-reptiele
Die twee nie-archosourus-reptiele, Testudines (skilpaaie) en Lepidosauria (slangee, akkedisse en wurmakkedisse), asook Choristodera (semi-waterdiere wat in die vroeë Mioseen uitgesterf het), het die uitwissing oorleef.[21] Meer as 80% van die Kryt se skilpaaie het oorleef. Daarbenewens het al ses skilpadfamilies wat aan die einde van die Kryt voorgekom het, oorleef tot in die Paleogeen en hulle word deur huidige spesies verteenwoordig.[47]
Die orde Squamata, wat vandag verteenwoordig word deur akkedisse, slange en wurmakkedisse, het verskeie ekologiese nisse gevul tydens die Jura en was suksesvol tydens die Kryt. Hulle het die uitwissing oorleef en is tans die suksesvolste en uiteenlopendse groep lewende reptiele met meer as 6 000 spesies. Nie ’n enkele familie van aardlewende Squamata is uitgewis nie en fossiele getuienis dui daarop dat hulle geen noemenswaardige afname in getalle beleef het nie. Hul klein grootte, aanpasbare metabolisme en vermoë om na gunstiger habitats te trek, was sleutelfaktore in die feit dat hulle oorleef het.[21][47] Reusagtige nie-Archosourus-waterreptiele soos Mosasauridae en Plesiosauria, wat die belangrikste mariene roofdiere van hul tyd was, het aan die einde van die Kryt uitgesterf.[48][49] Hulle was waarskynlik minder in staat om by veranderende toestande aan te pas weens hul grootte.
Archosourusse
Die archosourus-klade sluit twee huidige ordes in, Crocodilia (waarvan Alligatoridae, Crocodylidae en Gavialidae die enigste oorlewende families is) en dinosourusse (waarvan voëls die enigste oorlewende lede is), asook die uitgestorwe nie-vlieënde dinosourusse en die pterosourus.
Voëls
Die meeste paleontoloë beskou voëls as die enigste oorlewende dinosourusse. Alle nie-Neornithes-voëls het egter uitgesterf, insluitende florerende groepe soos Enantiornithes en Hesperornithes.[50] Verskeie ondersoeke van voëlfossiele toon die splitsing van spesies voor die K-Pg-grens en dat verwante van eende, hoenders en loopvoëls saam met nie-vlieënde dinosorusse bestaan het.[51] Groot versamelings voëlfossiele lewer voldoende bewys dat daar in die Kryt ’n groot verskeidenheid spesies was wat nie die Paleogeen gehaal het nie. ’n Baie klein deeltjie van Kryt se die voëls het dus oorleef en uit hulle het vandag se voëls ontwikkel.[52][53] Die enigste subklas voëls wat beslis die uitwissing oorleef het, is Neornithes (hoewel een Paleogeen-spesie, Qinornis paleocenica, verteenwoordigend kan wees van ’n nie-Neornithes-voël wat oorleef het.[52] Neornithes kon vermoedelik die uitwissing oorleef danksy hul vermoë om te kan duik, swem of skuiling soek in water en moeraslande. Baie spesies kan ook neste bou en in gate in bome of mierneste bly, wat hulle ook kon beskerm het. Hulle het ná die uitwissing ekologiese nisse gevul wat leeg gelaat is deur die uitsterwing van die nie-vlieënde dinosourusse.[54]
Nie-vlieënde dinosourusse

Buiten ’n paar omstrede bewerings, stem wetenskaplikes saam dat alle nie-vlieënde dinosourusse uitgesterf het by die K-Pg-grens. Die dinisourus-fossielrekord is geïnterpreteer as dat daar óf ’n afname óf geen afname in verskeidenheid was in die laaste paar miljoen jaar van die Kryt, en dit kan wees dat die gehalte van die fossielrekord net te swak was om tussen die twee opsies te onderskei.[55] Aangesien daar geen bewys is dat nie-vlieënde dinosourusse gate kon grawe, swem of duik nie, was hulle nie in staat om hulself te beskerm teen die omgewingstres wat by die K-Pg-grens voorgekom het nie. Kleiner diere (benewens voëls) kon dalk oorleef het, maar hulle sou geen kos gehad het nie aangesien plante skaars sou gewees het vir plantvretende diere en dit sou weer ’n voedseltekort veroorsaak het vir vleisvretende diere.[54]
Die toenemende konsensus oor die endotermie ("warmbloedigheid") van dinosourusse help om hul uitwissing te verstaan, terwyl hul verwante die krokodille oorleef het. Die ektotermiese ("koudbloedige") krokodille sou ’n klein behoefte aan kos gehad het aangesien hulle verskeie maande sonder kos kan klaarkom, terwyl endotermiese diere van dieselfde grootte meer kos nodig sou gehad het vir hul vinniger metabolisme. Nie-vlieënde dinosourusse sou dus uitgesterf het weens ’n tekort aan kos,[18] terwyl sommige krokodille oorleef het. Die oorlewing van ander warmbloedige diere, soos soogdiere, kon danksy ander redes gewees het soos hul kleiner behoefte aan kos vanweë hul kleiner grootte in dié tyd.[56]
Soogdiere
Alle belangrike soogdierlyne uit die Kryt, insluitende kloaakdiere (eierlêende soogdiere), Multituberculata, buidel- en plasentadiere, Dryolestoidea[57] en Gondwanatheria[58] het die uitwissing oorleef hoewel daar ook groot verliese was. Buideldiere het grootliks uit Noord-Amerika verdwyn, en die Asiatiese Deltatheroida, primitiewe verwante van bestaande buideldiere, is uitgewis.[59]
Die soogdiere wat die uitwissing oorleef het, was oor die algemeen klein – omtrent so groot soos rotte. Dit sou hulle gehelp het om skuiling te vind. Daar word ook gemeen baie van hulle kon gate grawe of was semi-waterdiere omdat daar vandag verskeie sulke soogdierspesies is. Dit sou bykomende beskutting gebied het teen omgewingstres.[54]
Chicxulub-asteroïde

Bewyse van ’n impak
In 1980 het ’n span navorsers bestaande uit die Nobelprys-wenner en fisikus Luis Alvarez, sy seun die geoloog Walter Alvarez en die chemici Frank Asaro en Helen Michel ontdek sedimentlae oor die hele wêreld heen bevat by die K-Pg-grens ’n konsentrasie iridium wat veel hoër is as normaal (30, 160 en 20 keer in dele wat oorspronklik bestudeer is). Iridium is uiters skaars in die aardkors, maar kom algemeen voor in die meeste asteroïdes en komete. Die Alvarez-span het voorgestel dat ’n asteroïde die Aarde getref het in die tyd van die K-Pg-grens.[3] Daar was vroeër gissings oor die moontlikheid van ’n impak, maar dit was die eerste konkrete bewys daarvan.[60]
Die teorie is eers as radikaal beskou, maar nog bewyse is kort daarna ontdek. Daar is gevind die grensklei is vol uiters klein bolletjies rots, wat gevorm is deur druppels gesmelte rots van die impak.[61] Misvormde kwarts en ander minerale is ook in die K-Pg-grens gevind.[62][63] Sulke misvormde minerale word veroorsaak deur intense druk soos met kernontploffings of ’n meteoriet-impak. Die ontdekking van neerslae wat deur ’n tsoenami veroorsaak is aan die kus van die Golf van Mexiko en die Karibiese See het ook bewys gelewer van ’n impak,[64] en het daarop gedui die impak kon daarnaby plaasgevind het – net soos die ontdekking dat die K-Pg-grens dikker word in die suid-VSA – tot meters dik in Noord-Nieu-Mexiko.[11]

Deur verdere navorsing is gevind die Chicxulub-krater onder Chicxulub aan die kus van Yucatán, Mexiko, is die bron van die K-Pg-grensklei. Die krater is ovaal, met ’n gemiddelde deursnee van rofweg 180 km – omtrent die grootte wat die Alvarez-span gereken het dit sou wees.[65] Dit het onteenseglik bewys dat die K-Pg-uitwissing deur ’n botsing veroorsaak is.
Uitwerking van ’n impak
So ’n groot impak sou fotosintese beperk het deur ’n stofwolk te veroorsaak wat die sonlig vir ’n jaar of minder sou blokkeer, en deur ’n sproei van swaelsuur die stratosfeer in te stuur. Dit sou sonlig verder verminder het – met 10 tot 20%. Dit sou minstens 10 jaar neem om te verdamp, wat die uitwissing van plante en fitoplankton verduidelik, asook van organismes wat van hulle afhanklik is (insluitende plantvreters en roofdiere). Klein diere waarvan die voedselketting op detritus (dooie organiese materiaal) berus, sou ’n redelike kans gehad het om te oorleef.[56]
Brande het ook waarskynlik wêreldwyd voorgekom weens die hitte en brandende brokstukke wat deur die impak veroorsaak is. Onlangse navorsing het getoon die wêreldwye puinlaag wat deur die impak gevorm is, bevat genoeg roet om te kan aflei die hele biosfeer het gebrand.[66] Die hoë O2-vlak tydens die laat-Kryt sou ontbranding aangehelp het. Die O2-vlak het in die vroeë Kanosoïkum skerp gedaal. As wydverspreide brande voorgekom het, sou dit die CO2-inhoud van die atmosfeer verhoog het en ’n tydelike kweekhuiseffek veroorsaak het sodra die stof gaan lê het, en dit sou die kwesbaarste organismes wat die tydperk net ná die impak oorleef het, uitgewis het.[67]
Die vorm en ligging van die krater dui ook aan dat groot verwoesting aangerig is benewens die stofwolk. Die asteroïde het in die see geland en sou enorme tsoenami's veroorsaak het, waarvoor bewyse gevind is op verskeie plekke in die Oos-VSA en om die Karibiese See – seesand op plekke wat toe in die binneland geleë was, en plantoorblyfsels en aardrotse in see-sedimente wat dateer uit die tyd van die impak.
Die meeste paleontoloë stem saam dat ’n asteroïde die Aarde na aan die einde van die Kryt-periode getref het, maar daar word gedebatteer of dit die enigste rede vir die uitwissing was.[68][69] Daar is bewyse van ’n interval van sowat 300 000 jaar vanaf die impak tot die massauitwissing.[70] In 1997 het die paleontoloog Sankar Chatterjee aandag gevestig op die veel groter Shiva-krater met ’n deursnee van 600 km, en op die moontlikheid van verskeie botsings.
In Maart 2010 het ’n internasionale paneel wetenskaplikes die asteroïde-teorie, spesifiek die Chicxulub-impak, onderskryf as die oorsaak van die uitwissing. ’n Span van 41 wetenskaplikes het 20 jaar se wetenskaplike geskrifte nagevors en enige teorieë oor enorme vulkaniese uitbarstings verwerp. Hulle het vasgestel die uitwissing is veroorsaak deur die Chicxulub-impak, wat soveel energie opgewek het as 100 teraton TNT – meer as ’n miljard keer die energie van die bomme op Nagasaki en Hirosjima.[5]
Verwysings
- ↑ Renne, Paul R. (7 Februarie 2013). "Time Scales of Critical Events Around the Cretaceous-Paleogene Boundary". Science. 339 (6120): 684–687. Bibcode:2013Sci...339..684R. doi:10.1126/science.1230492.
{{cite journal}}: Onbekende parameter|coauthors=geïgnoreer (hulp) - ↑ Fortey, R (1999). Life: A Natural History of the First Four Billion Years of Life on Earth. Vintage. pp. 238–260. ISBN 978-0-375-70261-7.
- ↑ 3,0 3,1 Alvarez LW, Alvarez W, Asaro F, Michel HV (1980). "Extraterrestrial cause for the Cretaceous–Tertiary extinction". Science. 208 (4448): 1095–1108. Bibcode:1980Sci...208.1095A. doi:10.1126/science.208.4448.1095.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) Verwysingfout: Invalid<ref>tag; name "Alvarez" defined multiple times with different content - ↑ Hildebrand, A. R., G. T. Penfield, et al. (1991). "Chicxulub crater: a possible Cretaceous/Tertiary boundary impact crater on the Yucatan peninsula, Mexico." Geology 19: 867-871.
- ↑ 5,0 5,1 5,2 Schulte, P.; Alegret, L.; Arenillas, I.; Arz, J. A.; Barton, P. J.; Bown, P. R.; Bralower, T. J.; Christeson, G. L.; Claeys, P.; Cockell, C. S.; Collins, G. S.; Deutsch, A.; Goldin, T. J.; Goto, K.; Grajales-Nishimura, J. M.; Grieve, R. A. F.; Gulick, S. P. S.; Johnson, K. R.; Kiessling, W.; Koeberl, C.; Kring, D. A.; MacLeod, K. G.; Matsui, T.; Melosh, J.; Montanari, A.; Morgan, J. V.; Neal, C. R.; Nichols, D. J.; Norris, R. D.; Pierazzo, E. (5 Maart 2010). "The Chicxulub Asteroid Impact and Mass Extinction at the Cretaceous- Paleogene Boundary |". Science 327 (5970): 1214–1218. Bibcode:2010Sci...327.1214S.doi:10.1126/science.1177265
- ↑ Keller, G. (2012). The Cretaceous--Tertiary Mass Extinction, Chicxulub Impact, and Deccan Volcanism. Earth and Life, Springer: 759--793.
- ↑ Longrich N. R., Tokaryk T. T.; et al. (2011). "Mass extinction of birds at the Cretaceous-Paleogene (K–Pg) boundary". Proceedings of the National Academy of Sciences. 108 (37): 15253–15257.
{{cite journal}}: Uitdruklike gebruik van et al. in:|author=(hulp) - ↑ 8,0 8,1 Longrich, N. R., A.-B. S. Bhullar, et al. (2012). "Mass extinction of lizards and snakes at the Cretaceous-Paleogene boundary." Proceedings of the National Academy of Sciences doi:10.1073/pnas.1211526110
- ↑ Labandeira, C. C., K. R. Johnson, et al. (2002). "Preliminary assessment of insect herbivory across the Cretaceous-Tertiary boundary: major extinction and minimum rebound." Geological Society of America Special Paper 361: 297-327.
- ↑ Rehan, Sandra M.; Leys, Remko; Schwarz, Michael P. (2013). "First Evidence for a Massive Extinction Event Affecting Bees Close to the K-T Boundary". PLoS ONE. 8 (10): e76683. doi:10.1371/journal.pone.0076683. ISSN 1932-6203.
- ↑ 11,0 11,1 11,2 11,3 11,4 11,5 11,6 Nichols, D. J. and K. R. Johnson (2008). Plants and the K–T Boundary. Cambridge, Cambridge University Press.
- ↑ Friedman M (2009). "Ecomorphological selectivity among marine teleost fishes during the end-Cretaceous extinction". PNAS. 106: 5218–5223.
- ↑ Jablonski, D., 1994. Extinctions in the fossil record (and discussion). Philosophical Transactions of the Royal Society of London, Series B. 344, 11-17.
- ↑ Alroy J (1999). "The fossil record of North American Mammals: evidence for a Palaeocene evolutionary radiation". Systematic Biology. 48 (1): 107–118.
- ↑ Feduccia A (1995). "Explosive evolution in Tertiary birds and mammals". Science. 267: 637–638.
- ↑ Friedman M (2010). "Explosive morphological diversification of spiny-finned teleost fishes in the aftermath of the end-Cretaceous extinction". Proceedings of the Royal Society B. 277: 1675–1683.
- ↑ Weishampel, D. B., P. M. Barrett, et al. (2004). Dinosaur Distribution. The Dinosauria. D. B. Weishampel, P. Dodson and H. Osmolska. Berkeley, University of California Press: 517-606.
- ↑ 18,0 18,1 Wilf P, Johnson KR (2004). "Land plant extinction at the end of the Cretaceous: a quantitative analysis of the North Dakota megafloral record". Paleobiology. 30 (3): 347–368. doi:10.1666/0094-8373(2004)030<0347:LPEATE>2.0.CO;2. ISSN 0094-8373.
- ↑ Pospichal JJ (1996). "Calcareous nannofossils and clastic sediments at the Cretaceous–Tertiary boundary, northeastern Mexico". Geology. 24 (3): 255–258. Bibcode:1996Geo....24..255P. doi:10.1130/0091-7613(1996)024<0255:CNACSA>2.3.CO;2.
- ↑ Bown P (2005). "Selective calcareous nannoplankton survivorship at the Cretaceous–Tertiary boundary". Geology. 33 (8): 653–656. Bibcode:2005Geo....33..653B. doi:10.1130/G21566.1.
- ↑ 21,0 21,1 21,2 21,3 21,4 21,5 21,6 21,7 21,8 MacLeod N, Rawson PF, Forey PL, Banner FT, Boudagher-Fadel MK, Bown PR, Burnett JA, Chambers, P, Culver S, Evans SE, Jeffery C, Kaminski MA, Lord AR, Milner AC, Milner AR, Morris N, Owen E, Rosen BR, Smith AB, Taylor PD, Urquhart E, Young JR (1997). "The Cretaceous–Tertiary biotic transition". Journal of the Geological Society. 154 (2): 265–292. doi:10.1144/gsjgs.154.2.0265.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Gedl P (2004). "Dinoflagellate cyst record of the deep-sea Cretaceous-Tertiary boundary at Uzgru, Carpathian Mountains, Czech Republic". Geological Society, London, Special Publications. 230: 257–273. Bibcode:2004GSLSP.230..257G. doi:10.1144/GSL.SP.2004.230.01.13.
- ↑ MacLeod N (1998). "Impacts and marine invertebrate extinctions". Geological Society, London, Special Publications. 140 (1): 217–246. Bibcode:1998GSLSP.140..217M. doi:10.1144/GSL.SP.1998.140.01.16.
- ↑ Courtillot, V (1999). Evolutionary Catastrophes: The Science of Mass Extinction. Cambridge University Press. p. 2. ISBN 0-521-58392-6.
- ↑ Glaessner MF (1937). "Studien über Foraminiferen aus der Kreide und dem Tertiär des Kaukasus: die Foraminiferen der ältesten Tertiärschichten des Nordwestkaukasus". Проблемс оф палеонтологий - Problems of Paleontology (in German). 2–3: 349–410. OCLC 776158910.
{{cite journal}}: AS1-onderhoud: onerkende taal (link) - ↑ Galeotti S, Bellagamba M, Kaminski MA, and Montanari A (2002). "Deep-sea benthic foraminiferal recolonisation following a volcaniclastic event in the lower Campanian of the Scaglia Rossa Formation (Umbria-Marche Basin, central Italy)" (PDF). Marine Micropaleontology. 44: 57–76. Besoek op 19 Augustus 2007.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Kuhnt W, Collins ES (1996). "8. Cretaceous to Paleogene benthic foraminifers from the Iberia abyssal plain" (PDF). Proceedings of the Ocean Drilling Program, Scientific Results. 149: 203–216. Besoek op 19 Augustus 2007.
- ↑ Vescsei A, Moussavian E (1997). "Paleocene reefs on the Maiella Platform Margin, Italy: An example of the effects of the cretaceous/tertiary boundary events on reefs and carbonate platforms". Facies. 36 (1): 123–139. doi:10.1007/BF02536880.
- ↑ Rosen BR, Turnšek D (1989). Jell A; Pickett JW (reds.). "Proceedings of the Fifth International Symposium on Fossil Cnidaria including Archaeocyatha and Spongiomorphs". Memoir of the Association of Australasian Paleontology. Brisbane, Queensland (8): 355–370.
{{cite journal}}:|contribution=ignored (hulp) - ↑ Ward PD, Kennedy WJ, MacLeod KG, Mount JF (1991). "Ammonite and inoceramid bivalve extinction patterns in Cretaceous/Tertiary boundary sections of the Biscay region (southwestern France, northern Spain)". Geology. 19 (12): 1181–1184. Bibcode:1991Geo....19.1181W. doi:10.1130/0091-7613(1991)019<1181:AAIBEP>2.3.CO;2.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Harries PJ, Johnson KR, Cobban WA, Nichols DJ (2002). "Marine Cretaceous-Tertiary boundary section in southwestern South Dakota: Comment and Reply". Geology. 30 (10): 954–955. Bibcode:2002Geo....30..954H. doi:10.1130/0091-7613(2002)030<0955:MCTBSI>2.0.CO;2. ISSN 0091-7613.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Vescsei A, Moussavian E (1997). "Paleocene reefs on the Maiella Platform Margin, Italy: An example of the effects of the cretaceous/tertiary boundary events on reefs and carbonate platforms". Facies. 36 (1): 123–139. doi:10.1007/BF02536880.
- ↑ Rosen BR, Turnšek D (1989). Jell A; Pickett JW (reds.). "Proceedings of the Fifth International Symposium on Fossil Cnidaria including Archaeocyatha and Spongiomorphs". Memoir of the Association of Australasian Paleontology. Brisbane, Queensland (8): 355–370.
{{cite journal}}:|contribution=ignored (hulp) - ↑ Patterson, C (1993). Osteichthyes: Teleostei. In: The Fossil Record 2 (Benton, MJ, editor). Springer. pp. 621–656. ISBN 0-412-39380-8.
- ↑ 35,0 35,1 Labandeira Conrad C, Johnson Kirk R, Wilf Peter (2002). "Impact of the terminal Cretaceous event on plant–insect associations" (PDF). Proceedings of the National Academy of Sciences of the United States of America. 99 (4): 2061–2066. Bibcode:2002PNAS...99.2061L. doi:10.1073/pnas.042492999. PMC 122319. PMID 11854501.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Wilf P, Labandeira CC, Johnson KR, Ellis B (2006). "Decoupled Plant and Insect Diversity After the End-Cretaceous Extinction". Science. 313 (5790): 1112–1115. Bibcode:2006Sci...313.1112W. doi:10.1126/science.1129569. PMID 16931760.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Vajda Vivi, Raine J Ian, Hollis Christopher J (2001). "Indication of Global Deforestation at the Cretaceous–Tertiary Boundary by New Zealand Fern Spike". Science. 294 (5547): 1700–1702. Bibcode:2001Sci...294.1700V. doi:10.1126/science.1064706. PMID 11721051.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Wilf P., Johnson K. R. (2004). "Land plant extinction at the end of the Cretaceous: a quantitative analysis of the North Dakota megafloral record". Paleobiology. 30 (3): 347–368.
- ↑ Johnson, KR (1991). Megafloral change across the Cretaceous Tertiary boundary in the northern Great Plains and Rocky Mountains. In: Global Catastrophes in Earth History: An Interdisciplinary Conference on Impacts, Volcanism, and Mass Mortality, Sharpton VI and Ward PD (redakteurs). Geological Society of America. ISBN 978-0-8137-2247-4.
{{cite book}}: Onbekende parameter|coauthors=geïgnoreer (hulp) - ↑ Askin, RA (1996). Palynological change across the Cretaceous–Tertiary boundary on Seymour Island, Antarctica: environmental and depositional factors. In: Cretaceous–Tertiary Mass Extinctions: Biotic and Environmental Changes, Keller G, MacLeod N (editors). WW Norton. ISBN 978-0-393-96657-2.
{{cite book}}: Onbekende parameter|coauthors=geïgnoreer (hulp) - ↑ Schultz PH, D'Hondt S (1996). "Cretaceous–Tertiary (Chicxulub) impact angle and its consequences". Geology. 24 (11): 963–967. Bibcode:1996Geo....24..963S. doi:10.1130/0091-7613(1996)024<0963:CTCIAA>2.3.CO;2. Besoek op 5 Julie 2007.
- ↑ Vajda V, McLoughlin S (2004). "Fungal Proliferation at the Cretaceous–Tertiary Boundary" (PDF). Science. 303 (5663): 1489–1490. doi:10.1126/science.1093807. PMID 15001770. Besoek op 7 Julie 2007.
- ↑ Fawcett, J. A.; Maere, S.; Van de Peer, Y. (April 2009). "Plants with double genomes might have had a better chance to survive the Cretaceous–Tertiary extinction event". Proceedings of the National Academy of Sciences of the United States of America 106 (14): 5737–5742.Bibcode:2009PNAS..106.5737F. doi:10.1073/pnas.0900906106.ISSN 0027-8424. PMC 2667025
- ↑ Archibald JD, Bryant LJ (1990). "Differential Cretaceous–Tertiary extinction of nonmarine vertebrates; evidence from northeastern Montana. In: Global Catastrophes in Earth History: an Interdisciplinary Conference on Impacts, Volcanism, and Mass Mortality (Sharpton VL and Ward PD, editors)". Geological Society of America, Special Paper. 247: 549–562.
- ↑ Estes, R. (1964). "Fossil vertebrates from the Late Cretaceous Lance Formation, Eastern Wyoming." University of California Publications, Department of Geological Sciences 49: 1-180.
- ↑ Gardner J. D. (2000). "Albanerpetontid amphibians from the Upper Cretaceous (Campanian and Maastrichtian) of North America". Geodiversitas. 22 (3): 349–388.
- ↑ 47,0 47,1 Novacek MJ (1999). "100 Million Years of Land Vertebrate Evolution: The Cretaceous-Early Tertiary Transition". Annals of the Missouri Botanical Garden. 86 (2): 230–258. doi:10.2307/2666178. JSTOR 2666178.
- ↑ Chatterjee S, Small BJ (1989). "New plesiosaurs from the Upper Cretaceous of Antarctica". Geological Society, London, Special Publications. 47 (1): 197–215. Bibcode:1989GSLSP..47..197C. doi:10.1144/GSL.SP.1989.047.01.15. Besoek op 4 Julie 2007.
- ↑ O'Keefe FR (2001). "A cladistic analysis and taxonomic revision of the Plesiosauria (Reptilia: Sauropterygia)". Acta Zoologica Fennica. 213: 1–63.
- ↑ Hou L, Martin M, Zhou Z, Feduccia A (1996). "Early Adaptive Radiation of Birds: Evidence from Fossils from Northeastern China". Science. 274 (5290): 1164–1167. Bibcode:1996Sci...274.1164H. doi:10.1126/science.274.5290.1164. PMID 8895459.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Clarke JA, Tambussi CP, Noriega JI, Erickson GM, Ketcham RA (2005). "Definitive fossil evidence for the extant avian radiation in the Cretaceous". Nature. 433 (7023): 305–308. Bibcode:2005Natur.433..305C. doi:10.1038/nature03150. PMID 15662422.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ 52,0 52,1 Longrich, Nicholas R.; Tokaryk, Tim; Field, Daniel J. (2011). "Mass extinction of birds at the Cretaceous–Paleogene (K–Pg) boundary". Proceedings of the National Academy of Sciences. 108 (37): 15253–15257. Bibcode:2011PNAS..10815253L. doi:10.1073/pnas.1110395108.
- ↑ "Primitive Birds Shared Dinosaurs' Fate". Science Daily. 20 September 2011. Besoek op 20 September 2011.
- ↑ 54,0 54,1 54,2 Robertson DS, McKenna MC, Toon OB, Hope S, Lillegraven JA (2004). "Survival in the first hours of the Cenozoic" (PDF). GSA Bulletin. 116 (5–6): 760–768. Bibcode:2004GSAB..116..760R. doi:10.1130/B25402.1. Besoek op 31 Augustus 2007.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ David, Archibald (2004). "Dinosaur Extinction". In Weishampel David B, Dodson Peter, Osmólska Halszka (eds.) (red.). The Dinosauria (2nd uitg.). Berkeley: University of California Press. pp. 672–684. ISBN 0-520-24209-2.
{{cite book}}:|editor=has generic name (hulp); Onbekende parameter|chapterurl=geïgnoreer (hulp); Onbekende parameter|coauthors=geïgnoreer (hulp)AS1-onderhoud: meer as een naam (link) - ↑ 56,0 56,1 Ocampo, A (2006). "Unravelling the Cretaceous–Paleogene (K–T) turnover, evidence from flora, fauna and geology in biological processes associated with impact events". In Cockell C, Gilmour I, Koeberl C (red.). Biological Processes Associated with Impact Events. SpringerLink. pp. 197–219. ISBN 978-3-540-25735-6.
{{cite book}}: Onbekende parameter|coauthors=geïgnoreer (hulp)AS1-onderhoud: meer as een naam (link) - ↑ Gelfo JN and Pascual R (2001). "Peligrotherium tropicalis (Mammalia, Dryolestida) from the early Paleocene of Patagonia, a survival from a Mesozoic Gondwanan radiation" (PDF). Geodiversitas. 23: 369–379.
- ↑ Goin FJ, Reguero MA, Pascual R, von Koenigswald W, Woodburne MO, Case JA, Marenssi SA, Vieytes C, Vizcaíno SF (2006). "First gondwanatherian mammal from Antarctica". Geological Society, London, Special Publications. 258: 135–144. Bibcode:2006GSLSP.258..135G. doi:10.1144/GSL.SP.2006.258.01.10.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ McKenna, MC (1997). Classification of mammals: above the species level. Columbia University Press. ISBN 978-0-231-11012-9.
{{cite book}}: Onbekende parameter|coauthors=geïgnoreer (hulp) - ↑ De Laubenfels MW (1956). "Dinosaur extinction: One more hypothesis". Journal of Paleontology. 30 (1): 207–218. JSTOR 1300393.
- ↑ Smit J., Klaver J. (1981). "Sanidine spherules at the Cretaceous-Tertiary boundary indicate a large impact event". Nature. 292: 47–49.
- ↑ Bohor B. F., Foord E. E.; et al. (1984). "Mineralogic evidence for an impact event at the Cretaceous-Tertiary boundary". Science. 867: 869.
{{cite journal}}: Uitdruklike gebruik van et al. in:|author=(hulp) - ↑ Bohor B. F., Modreski P. J.; et al. (1987). "Shocked Quartz in the Cretaceous-Tertiary Boundary Clays: Evidence for a Global Distribution". Science. 236 (4802): 705–709.
{{cite journal}}: Uitdruklike gebruik van et al. in:|author=(hulp) - ↑ Bourgeois J., Hansen T. A.; et al. (1988). "A tsunami deposit at the Cretaceous-Tertiary boundary in Texas". Science. 241 (4865): 567–570.
{{cite journal}}: Uitdruklike gebruik van et al. in:|author=(hulp) - ↑ Pope KO, Ocampo AC, Kinsland GL, Smith R (1996). "Surface expression of the Chicxulub crater". Geology. 24 (6): 527–530. Bibcode:1996Geo....24..527P. doi:10.1130/0091-7613(1996)024<0527:SEOTCC>2.3.CO;2. PMID 11539331.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Robertson, D.S., Lewis, W.M., Sheehan, P.M. & Toon, O.B. (2013) K/Pg Extinction: Re-evaluation of the Heat/Fire Hypothesis.Journal of Geophysical Research: Biogeosciences
- ↑ Pope KO, Baines KH, Ocampo AC, Ivanov BA (1997). "Energy, volatile production, and climatic effects of the Chicxulub Cretaceous/Tertiary impact". Journal of Geophysical Research. 102 (E9): 21645–21664. Bibcode:1997JGR...10221645P. doi:10.1029/97JE01743. PMID 11541145.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Keller G, Adatte T, Stinnesbeck W, Rebolledo-Vieyra, Fucugauchi JU, Kramar U, Stüben D (2004). "Chicxulub impact predates the K–T boundary mass extinction". PNAS. 101 (11): 3753–3758. Bibcode:2004PNAS..101.3753K. doi:10.1073/pnas.0400396101. PMC 374316. PMID 15004276.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Morgan J, Lana C, Kersley A, Coles B, Belcher C, Montanari S, Diaz-Martinez E, Barbosa A, Neumann V (2006). "Analyses of shocked quartz at the global K-P boundary indicate an origin from a single, high-angle, oblique impact at Chicxulub". Earth and Planetary Science Letters. 251 (3–4): 264–279. Bibcode:2006E&PSL.251..264M. doi:10.1016/j.epsl.2006.09.009.
{{cite journal}}: AS1-onderhoud: meer as een naam (link) - ↑ Keller G, Abramovich S, Berner Z, Adatte T (1 Januarie 2009). "Biotic effects of the Chicxulub impact, K–Pg catastrophe and sea level change in Texas". Palaeogeography, Palaeoclimatology, Palaeoecology. 271 (1–2): 52–68. doi:10.1016/j.palaeo.2008.09.007.
{{cite journal}}: AS1-onderhoud: meer as een naam (link)
Eksterne skakels
- "The Great Chicxulub Debate 2004". Geological Society of London. 2004. Besoek op 2 Augustus 2007.
- "The Chicxulub Debate". Princeton University. 2007. Besoek op 2 Augustus 2007.
- McLean D (1995). "The Deccan Traps Volcanism-Greenhouse Dinosaur Extinction Theory". Universiteit van Vermont. Besoek op 2 Augustus 2007.
- "What killed the dinosaurs?". University of California Museum of Paleontology. 1995. Besoek op 2 Augustus 2007.
 Wikimedia Commons het meer media in die kategorie Kryt-Paleogeen-uitwissing.
Wikimedia Commons het meer media in die kategorie Kryt-Paleogeen-uitwissing.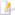 Hierdie artikel is vertaal uit die Engelse Wikipedia
Hierdie artikel is vertaal uit die Engelse Wikipedia